第七章 能量代谢和体温
第一节 能量代谢
新陈代谢是机体生命活动的基本特征,新陈代谢包括物质代谢与相传伴的能量代谢,简称代谢。
糖、脂肪、蛋白质三种营养物质,经消化转变成为可吸收的小分子营养物质而被吸收入血。在细胞中,这些营养物质经过同化作用(合成代谢),构筑机体的组成成分或更新衰老的组织;同时经过异化作用(分解代谢)分解为代谢产物。合成代谢和分解代谢是物质代谢过程中互相联系的、不可分割的两个侧面。
在分解代谢过程中,营养物质蕴藏的化学能便释放出来。这些化学能经过转化,便成了机体各种生命活动的能源,所以说分解是代谢的放能反应。而在合成代谢过程中,需要供给能量,因此是吸能反应。可见,在物质代谢过程中,物质的变化与能量的代谢是紧密联系着的。生物体内物质代谢过程中所伴随的能量释放、转移和利用等,称为能量代谢(energy metabolism)。
机体所需的能量来源于食物中的糖、脂肪和蛋白质。这些能源物质分子结构中的碳氢键蕴藏着化学能,在氧化过程中碳氢键断裂,生成CO2和 H2O,同时释放出蕴藏的能。这些能量的50%以上迅速转化为热能,用于维持体温,并向体外散发。其余不足50%则以高能磷酸键的形式贮存于体内,供机体利用。体内最主要的高能磷酸键化学物是三磷酸腺苷( ATP)。此外,还可有高能硫酯键等。机体利用ATP去合成各种细胞组成分子、各种生物活性物质和其他一些物质;细胞利用ATP去进行各种离子和其它一些物质的主动转运,维持细胞两侧离子浓度差所形成的势能;肌肉还可利用ATP所载荷的自由能进行收缩和舒张,完成多种机械功。总的看来,除骨骼肌运动时所完成的机械功(外功)以外,其余的能量最后都转变为热能。例如心肌收缩所产生的势能(动脉血压)与动能(血液流速),均于血液在血管内流动过程中,因克服血流内、外所产生的阻力而转化为热能。在人体内,热能是最“低级”形式的能,热能不能转化为其它形式的能,不能用来作功。
本节主要叙述整个机体的能量代谢测定的原理与方法,基础代谢以及机体在某些状态下的代谢等问题,不涉及能量代谢的各个方面。
通用的法定计量的热量单位为焦耳(Joules.J)。过去热量单位是卡或千卡,1卡=4.187焦耳,1千卡=4.187千焦耳(kJ)。1焦耳/s为1瓦特。
一、能量代谢测定的原理和方法
热力学第一定律指出:能量由一种形式转化为另一种形式的过程中,既不能增加,也不减少。这是所有形式的能量(动能、热能、电能入化学能)互相转化的一般规律,也就是能量守恒定律。机体的能量代谢也遵循这一规律,即在整个能量转化过程中,机体所利用的蕴藏于食物中的化学能与最终转化成的热能和所作的外功,按能量来折算是完全相等的。因此,测定在一定时间内机体所消耗的食物,或者测定机体所产生的热量与所做的外功,都可测算出整个机体的能量代谢率(单位时间内所消耗的能量)。
测定整个机体单位时间内发散的总热量,通常有两类方法:直接测热法和间接测热法。
(一)直接测热法
直接测热法(direct calormetry)是测定整个机体在单位时间内向外界环境发散的总热量。此总热量就是能量代谢率。如果在测定时间内做一定的外功,应将外功(机械功)折算为热量一并计入。图7-1是本世纪初Arwater-Benedict所设计的呼吸热量计的结构模式图。它是由隔热密封的房间,其中设一个铜制的受试者居室。用调节温度的装置控制隔热壁与居室之间空气的温度,使之与居室内的温度相等,以防居室内的热量因传导而丧失。这样,受试者机体所散发的大部分热量便被居室内管道中流动的水所吸收。根据流过管道的水量和温度差,将水的比热考虑在内,就可测出水所吸收的热量。当然,受试者发散的热量有一部分包含在不感蒸发(参看第二节)量中,这在计算时也要加进去。受试者呼吸的空气由进出居室的气泵管道系统来供给。此系统中装有硫酸和钠石灰,用业吸收水蒸气和CO2。管道系统中空气中的O2则由氧气筒定时补给。
直接测热法的设备复杂,操作繁锁,使用不便,因而极少应用。一般都采用间接测热法。

图7-1 直接测热装置示意图
(二)间接测热法
在一般化学反应中,反应物的量与产物量之间呈一定的比例关系,这就是定比定律。例如,氧化1mol葡萄糖,需要6mol氧,同时产生6molCO2和6molH2O,并释放一定量的能。下列反应式表明了这种关系:
C6H12O6+602→6CO2+6H20+△H
同一种化学反应,不论经过什么样的中间步骤,也不论反应条件差异多大,这种定比关系仍然不变。例如,在人本内氧化1mol葡萄糖,同在体外氧化燃烧1mol葡萄糖一样,都要消耗6molCO2和6molH20,而且产生的热量也相等。一般化学反应的这种基本规律也见于人体内营养物质氧化供能的反应(蛋白质的情况下有些出入,参看下文),所以它成了能量代谢间接测热法的重要依据。
间接测热法(indirect calorimetry)的基本原理就是利用这种定比关系,查出一定时间内整个人体中氧化分解的糖、脂肪、蛋白质各有多少,然后据此计算出该段时间内整个机体所释放出来的热量。因此,必须解决两个问题:一是每种营养物质氧化分解时产生的能量有多少(即食物的热价);二要分清三种营养物质各氧化了多少。
食物的热价应用弹式热量计,在体外测定了一定量的的糖、脂肪和蛋白质燃烧时所释放的热量,并同这三类物质在动物体内氧化到最终产物C02和水时所产生的热量相比较,证明了糖和脂肪在体外燃烧与在体内氧化分解所产生的热量是相等的。于是将1g食物氧化(或在体外燃烧)时所释放出来的能量称为食物的热价(thermal equivalent of food)。食物的热价分为物理热价和生物热价。前者指食物在体外燃烧时释放的热量,后者系食物经过生物氧化所产生的热量。糖(或脂肪)的物理热价和生物热价是相等的,而蛋白质的生物热价则小于它的物理热价。因为蛋白质在体内不能被彻底氧化分解,它有一部分主要以尿素的形式从尿中排泄的缘故。三种营养物质在物理热价和生物热价见表演7-1。
呼吸商 机体依靠呼吸功能从外界摄取氧,以供各种营养物质氧化分解的需要,同时也将代谢终生物CO2呼出体外,一定时间内机体的CO2产量与耗氧量的比值称为呼吸商(respiratory quotient, RQ)。各种营养物质在细胞内氧化供能属于细胞呼吸过程,因而双将各种营养物质氧化时的CO2产量与耗氧量的比值称为某物质的呼吸商。严格说来,应该以CO2和O2的克分子(mol)比值来表示呼吸商。但是,因为在同一温度和气压条件下,容积相等的不同气体,其分子数都是相等的,所以通常都用容积数(ml或L)来计算CO2与O2的比值,即:
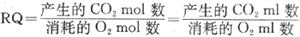
糖、脂肪和蛋白质氧化时,它们的CO2产量与耗氧量各不相同,三者的呼吸商也不一样。
因为各种营养物质无论在体内或体外氧化,它们的耗氧量与CO2产量都取决于各该物质的化学组成,所以,在理论上任何一种营养物质的呼吸商都可以根据它的氧化成终产物(CO2和H20)化学反应式计算出来的。
糖的一般分子式为(CH20)n,氧化时消耗的02和产生的C02分子数相等,呼吸商应该等于1。如上述葡萄糖氧化的反应式所示,C02产量与耗氧量均为6mol故:
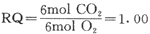
脂肪氧化时需要消耗更多的氧。在脂肪本身的分子结构中,氧的含量远较碳和氢少。因此,另外提供的氧不仅要用氧化脂肪分子中的碳,还要用来氧化其中的氢。所以脂肪的呼吸商将小于1。现以甘油三酸酯(triolein)为例:
RQ=57molC02/80mol02=0.71
蛋白质的呼吸商较难测算,因为蛋白质在体内不能完全氧化,而且它氧化分解途径的细节,有些还不够清楚,所以只能通过蛋白质分子中的碳和氢被氧化时势需氧量和C02产量,间接算出蛋白质的呼吸商,其计算值为0.80。
在人的日常生活中,营养物质不是单纯的,而是糖、脂肪和蛋白质混合而成的(混合膳食)。所以,呼吸商常变动于0.71-1.00之间。人体在特定时间内的呼吸产要看哪种营养物质是当时的主要能量来源而定。若能源主要是糖类,则呼吸商接近于1.00;若主要是脂肪,则呼吸商接近于0.71。在长期病理性饥饿情况下,能源主要来自机体本身的蛋白质和脂肪,则呼吸商接近于0.80。一般情况下,摄取混合食物时,呼吸商常在0.85左右。
现将糖、脂肪和蛋白质三者的热价、氧热窬及呼吸商等数据列于表7-1,以供理解和测算能量代谢率之用。
表7-1 三种营养物质氧化时的几种数据
| 营养
物质 |
产热量(kJ/g) |
耗氧量 |
CO2产量 |
氧热价 |
呼吸商 |
| 物理热价(用弹式热量计测得) |
生物热价(体内生物氧化什) |
营养学
热价* |
| (L/g) |
(L/g) |
(kJ/J) |
(RQ) |
| 糖 |
17 |
17 |
16.7 |
0.83 |
0.83 |
21 |
1.00 |
| 蛋白质 |
23.5 |
18 |
16.7 |
0.95 |
0.76 |
18.8 |
0.80 |
| 脂肪 |
39.8 |
39.8 |
37.7 |
2.03 |
1.43 |
19.7 |
0.71 |
*营养学通常采用概数来计算食物的热价
影响呼吸商的其它因素:
机体的组织、细胞 不仅能同时氧化分解各种营养物质,而且也使一种营养物质转变为另一种营养物质。糖的转化为脂肪时,呼吸商可能变大,甚至超过1.00。这是由于当一部分糖转化为脂肪时,原来糖分子中的氧即有剩余,这些氧可能参加机体代谢过程中氧化反应,相应地减少了从外界摄取的氧量,因而呼吸商变大。反过来,如果脂肪转化为糖,呼吸商也可能低于0.71。这是由于脂肪分子中含氧比例小,当转化为糖时,需要更多的氧进入分子结构,因而机体摄取并消耗外界氧的量增多,结果呼吸商变小。另外,还有其它一些代谢反应也能影响呼吸商。例如,肌肉剧烈运动时,由于氧供不应求,糖酵解增多,将有大量乳酸进入血液。乳酸和碳酸盐作用的结果,会有大量由肺肺排出,此时呼吸商将变大。又如,肺过度通气、酸中毒等情况下,机体中与生物氧化无关的CO2大量排出,也可现呼吸大于1.00的情况。相反,肺通气不足、碱中毒等情况下,呼吸商将降低。
前已述,应该测出在一定时间内机体中糖、脂肪和蛋白质三者氧化分解的比例。为此。首先必须查清氧化了多少蛋白质,并且将氧化这些蛋白质所消耗的氧量和所产生的CO2从机体在该时间内的总耗氧量和总CO2产量中减去,算出糖和脂肪氧化(非蛋白质代谢)的CO2产量和耗氧量的比值,即非蛋白呼吸商(non-protein respiratory quotient,NPRQ),然后才有可能进一步查清糖和脂肪各氧化了多少克。
尿氮测定 尿中的氮物质主要是蛋白质的分解产物。因此可以通过尿氮来估算体内被氧化的蛋白质的数量。蛋白质的平均重量组成是:C50%,O223%,N16%,S1%。蛋白质中16%的N是完全随尿排出的。所以,1g 尿氮相当于氧分解6.25g蛋白质,测得的尿氮重量(g)乘以6.25,便相当于体内氧分解的蛋白质量。
非蛋白呼吸商 它是估算非蛋白代谢中糖和脂肪氧化的相对数量的依据。研究工作者早已按从0.707到1.00范围内的非蛋白呼吸产,算出糖和脂肪两者氧化的各自百分比以及氧热价(表7-2)。
表7-2 非蛋白呼吸商和氧热价
| 非蛋白呼吸商 |
氧 化 的 % |
氧热价(kJ/L) |
| 糖 |
脂肪 |
| 0.707 |
0.00 |
100.0 |
19.62 |
| 0.71 |
1.10 |
98.9 |
19.64 |
| 0.72 |
4.75 |
95.2 |
19.69 |
| 0.73 |
8.40 |
91.6 |
19.74 |
| 0.74 |
12.0 |
88.0 |
19.79 |
| 0.75 |
15.6 |
84.4 |
19.84 |
| 0.76 |
19.2 |
80.8 |
19.89 |
| 0.77 |
22.8 |
77.2 |
19.95 |
| 0.78 |
26.3 |
73.7 |
19.99 |
| 0.79 |
29.0 |
70.1 |
20.05 |
| 0.80 |
33.4 |
66.6 |
20.10 |
| 0.81 |
36.9 |
63.1 |
20.15 |
| 0.82 |
40.3 |
59.7 |
20.20 |
| 0.83 |
43.8 |
56.2 |
20.26 |
| 0.84 |
47,2 |
52.8 |
20.31 |
| 0.85 |
50.7 |
49.3 |
20.36 |
| 0.86 |
54.1 |
45.9 |
20.41 |
| 0.87 |
57.5 |
42.5 |
20.46 |
| 0.88 |
60.8 |
39.2 |
20.51 |
| 0.89 |
64.2 |
35.8 |
20.56 |
| 0.90 |
67.5 |
32.5 |
20.61 |
| 0.91 |
70.8 |
29.2 |
20.67 |
| 0.92 |
74.1 |
25.9 |
20.71 |
| 0.93 |
77.4 |
22.6 |
20.77 |
| 0.94 |
80.7 |
19.3 |
20.82 |
| 0.95 |
84.0 |
16.0 |
20.87 |
| 0.96 |
87.2 |
12.8 |
20.93 |
| 0.97 |
90.4 |
9.58 |
20.98 |
| 0.98 |
93.6 |
6.37 |
21.03 |
| 0.99 |
96.8 |
3.18 |
21.08 |
| 1.00 |
100.0 |
0.0 |
21.13 |
Lusk修订
间接测热法计算原则 实验测得的机体24小时内的耗氧量和CO2产量以及尿氮量,根据表7-1和7-2中相应的一些数据计算。首先,由尿氮量算出被氧分解的蛋白质量。由被氧化的蛋白质量从表7-1中算出其产热量、耗氧量和CO2产量;其次从总耗氧量和总CO2产量中减去蛋白质耗氧量和CO2产量,计算出非蛋白呼吸商。根据非蛋白呼吸商查表7-2的相应的非蛋白呼吸商的氧热价,计算出非蛋白代谢的产热量;最后,24小时产热量为蛋白质代谢的产热量与非蛋白代谢的产热量之和。此外,从非蛋白呼吸还可推算出参加代谢的糖和脂肪的比例。
间接测热法的计算方法举例
首先测定受试者一定时间内的耗氧量和CO2产量,假定受试者24小时的耗氧量为400L,CO2产量为340L(已换算成标准状态的气体容积)。另经测定尿氮排出量为12g。根据这些数据和查表7-1、7-2,计算24小时产热量,其步骤如下:(1)蛋白质氧化量=12×6.25=7g
产热量=18×75=1350kJ
耗氧量=0.95×75=71.25L
CO2产量=0.76×75=57L
(2)非蛋白呼吸商
非蛋白代谢耗氧量=400-71.25=328.75L
非蛋白代谢CO2产量=340-57=283L
非蛋白呼吸商=283/328.75=0.86
(3)根据非蛋白呼吸商的氧热价计算非蛋白代谢的热量
查表7-2,非蛋白呼吸商为0.86时,氧热价为20.41。所以,非蛋白代谢产热量=328.75×20.41=6709.8kj 。
(4)计算24小时产热量
24小时产热量=1350+6709.8=8059.8kJ
(蛋白质代(非蛋白代谢产热量)谢产热量)
计算的最后数值8059.8kJ就是该受试者24小时内的能量代谢率
耗氧量与CO2产量的测定方法及临床应用 测定耗氧量和CO2产量的方法有两种:闭合式测定法和开放式测定法。
(1)闭合式测定法:在动物实验中,将受试动物置于一个密闭的能吸热的装置中。通过气泵,不断将定量的氧气送入装置。动物不断地摄取氧,可根据装置中氧量的减少计算出该动物在单位时间内的耗氧量。动物呼出的CO2则由装在气体回路中的CO2吸收剂吸收。然后根据实验前后CO2吸收剂的重量差,算出单位时间内的CO2产量。由耗氧量和CO2产量算出呼吸商。
临床上为了简便,通常只使用肺量计(图7-2)来测量耗氧量。该装置的气体中容器中装置氧气,受试者通过呼吸口瓣将氧气吸入呼吸器官。此时气体容器的上盖随吸气而下降,并由连于上盖的描笔记录在记录纸上。根据记录纸上的方格还可读出潮气量值。受试者的呼出气则通过吸收容器(呼出气中的CO2和水可除除掉)进入气体容器中,于是气体容器的上盖又复升高,描笔也了随之升高。由于受试者摄取了一定量的氧气,呼出气中CO2又被除掉,气体容器中的氧气量因而逐渐减少。描笔则记录出曲线逐渐下降的过程。在一定时间内(通常为6min),描笔的总下降高度,就是该时间内的耗氧量。
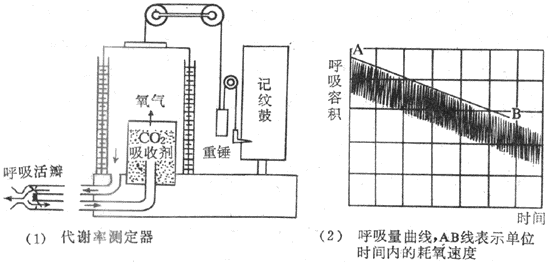
图7-2 肺量计结构模式图
(2)开放式测定法(气体分析法):它是在机体呼吸空气的条件下测定耗氧量和CO2产量的方法,所以称为开放法。其原理是,采集受试者一定时间内的呼出气,测定呼出气量并分析呼出气中氧和CO2的容积百分比。由于吸入气就是空气,所以其中氧和CO2的容积百分比不必另测。根据吸入气和呼出气中氧和CO2的容积百分比的差数,可算出该时间内的耗氧量和CO2排出量。
气体分析方法很多,最简便而又广泛应用的方法,是将受试者在一定时间内呼出气采集于气袋中,通过气量计测定呼气量,然后用气体分析器分析呼出气的组成成分,进而计算耗氧量和CO2产量,并算出呼吸商。
现举出一个气体分析实验例。
现某健康成人安静状态下的呼出气作气体分析,结果为:O2=16.26%;CO2=4.14%。呼出气量为1分钟5.2L(通常将呼出气量换算为不含水蒸气的标准状态值;也有换算为被水蒸气饱和的1个大气压、体温状态下的值)。空气的组成是:O2=20.96%;N=79.00%;CO2=0.04%。则:
受试者从每100ml通过肺的气体中吸收的氧气为:
20.96ml-16.26ml=4.7ml(或每升气体中的47ml)
(空气) (呼出气)
因为呼出气量为5.2L,则1分钟的耗氧量为:
47ml×5.2=244.4ml
100ml呼气中的CO2为:
4.14ml-0.04ml=4.1ml(或每升气体中的41ml)
(呼出气) (空气)
1分钟CO2排出量为:
41ml×5.2=213.2ml
RQ=213.1/244=0.85
所得呼吸商为受试者在安静状态下混合膳食代谢的呼吸商。
临床上和劳动卫生常采用简略法,即用气体分析法测得一定时间内的耗氧量和CO2产量,并求出呼吸商,并且不考虑蛋白质代谢部分,就根据非蛋白呼吸商表7-2查出呼吸商的氧热价,然后将氧热价乘以耗氧量,便得出该时间内的产热量。仍以上述间接测热法计算方法列举的例子为数据(见前),按此简略法来计算,则结果如下:
呼吸商=340/400=0.85
查表7-2,呼吸商0.85时的氧热价为20.36kJ,所以24小时的产热量=20.36×400=8144kJ。
这个数值与按完整的间接法计算得出的数值8059.8kJ是很近似的,误差都在1%-2%以下。而且在非蛋白呼吸商从0.70到1.00的范围内,氧热价也过变动于19.6-21.1之间。此法在实际工作中是可用的。
另一种更简便的简略法只利用肺量计测出受试者一定时间内(通常为6min)的耗氧量。受试者一般都吃混合膳食,所以通常将非蛋白呼吸商定为0.82,氧热价为20.20kJ。因此,测出一定时间内的耗氧量后,使可依下式来计算:
产热量=20.20×耗氧量(kJ)
二、影响能量代谢的因素
影响能量代谢的因素有肌肉活动、精神活动、食物的特殊动力作用和环境温度等。
(一)肌肉活动
肌肉活动对能量代谢的影响最为显著。机体任何轻微的活动都可提高代谢率。人在运动或劳动时耗量显著增加,因为肌肉活动需要补给能量,而能量则来自大量营养物质的氧化,导致机体耗氧量的增加。机体耗氧量的增加与肌肉活动的强度呈正比关系,耗氧量最多右达安静时的10-20倍。肌肉活动的强度称为肌肉工作的强度,也就是劳动强度。劳动强度通常用单位时间内机体的产热量来表示,也就是说,可以把能量代谢率作为评估劳动强度的指标。从表7-3可以看出劳动强度或运动时的能量代谢率的增长情况。
表7-3 运动或劳动时的能量代谢率
| 机体的状态 |
产 热 量 |
| 平均(kJ/m2・min) |
| 躺卧 |
2.73 |
| 开会 |
3.40 |
| 擦窗子 |
8.30 |
| 洗衣 |
9.98 |
| 扫地 |
11.37 |
| 打排球 |
17.05 |
| 打篮球 |
24.22 |
| 踢足球 |
24.98 |
(二)精神活动
脑的重量只占体重的2%,但在安静状态下,却有15%左右的循环血量进入脑循环系统,这说明脑组织的代谢水平是很高的。据测定。在安静状态下,100g脑组织的耗氧量为3.5ml/min(氧化的葡萄糖量为4.5mg/min),此值接近安静肌肉组织耗氧量的20倍,脑组织的代谢率虽然如此之高,但据测定,在睡眠中和在活跃的精神活动情况下,脑中葡萄糖的代谢率却几乎没有差异。可见,在精神活动中,中枢神经系统本身的代谢率即使有些增强,其程度也是可以忽略的。
人在平静地思考问题时,能量代谢受到的影响并不大,产热量增加一般不超过4%。但在精神处于紧张状态,如烦恼、恐惧或强烈情绪激动时,由于随之出现的无意识的肌紧张以及刺激代谢的激素释放增多等原因,产热量可以显著增加。因此,在测定基础代谢率时,受试者必须摒除精神紧张的影响。
(三)食物的特殊动力作用
在安静状态下摄入食物后,人体释放的热量比摄入的食物本身氧化后所产生的热量要多。例如摄入能产100kJ热量的蛋白质后,人体实际产热量为130kJ,额外多产生了30kJ热量,表明进食蛋白质后,机体产热量超过了蛋白质氧化后产热量的30%。食物能使机体产生“额外”热量的现象称为食物的特殊动力作用(specific dynamic action)。糖类或脂肪的食物特殊动力作用为其产热量的4%-6%,即进食能产100kJ热量的糖类或脂肪后,机体产热量为104-106kJ。而混合食物可使产热量增加10%左右。这种额外增加的热量不能被利用来作功,只能用于维持体温。因此,为了补充体内额外的热量消耗,机体必须多进食一些食物补充这份多消耗的能量。
食物特殊动力作用的机制尚未完全了解。这种现象在进食后1h左右开始,并延续到7-8h。有人将氨基酸注入静脉内,可出现与经口给予相同的代谢率增值现象,这些事实使人们推想,食后的“额外”热量可能来源于肝处理蛋白质分解产物时“额外”消耗的能量。因此,有人认为肝在接脱氨基反应中消耗了能量可能是“额外”热量产生的原因。
(四)环境温度
人(裸体或只着薄衣)安静时的能量代谢,在20-30℃的环境中最为稳定。实验证明,当环境温度低于20℃时,代谢率开始有所增加,在10℃以下,代谢率便显著增加。环境温度低时代谢率增加,主要是由于寒冷刺激反射地引起寒战以及肌肉紧张增强所致。在20-30℃时代谢稳定,主要是由于肌肉松驰的结果。当环境温度为30-45℃时,代谢率又会逐渐增加。这可能是因为体内化学过程的反应速度有所增加的缘故,这时还有发汗功能旺盛及呼吸、循环功能增强等因素的作用。
三、基 础 代 谢
基础代谢(basal metabolism)是指基础状态下的能量代谢。基础代谢率(basal metabolic rate,BMR)是指单位时间内的基础代谢,即在基础状态下,单位时间内的能量代谢。所谓基础状态是指人体处在清醒而又非常安静、不受肌肉活动、环境温度、食物及精神紧张等因素的影响时的状态。测定基础代谢率,要在清晨末进餐以前(即食后12-14h)进行。前一日晚餐最好是清淡菜肴,而且不要吃得太饱,这样,过了12-14h ,胃肠的消化和吸收活动已基本完毕,也排除了食物的特殊动力作用的影响。测定之前不应做剧烈的活动,而且必须静卧半小时以上。测定时平卧,全身肌肉要松驰,尽量排除肌肉活动的影响。这时还应要求受试者排除精神紧张的影响,如摒除焦虑、烦恼、恐惧等心理活动。室温要保持在20-25℃之间,以排除环境温度的影响。基本条件下的代谢率,比一般安静时的代谢率可低些(比清醒安静时低8%-10%)。基础代谢率以每小时,每平方米体表面积的产热量为单位,通常以kJ/m2・h来表示。要用每平方米体表面积而不用每公斤体重的产热量来表示,是因为基础代谢率的高低与体重并不成比例关系,而与体表面积基本上成正比。
表7-4 人体产热量和体表面积的关系
| 测定人数 |
平均体重(kg) |
体重范围 |
kJ/m2・24h |
| 6 |
48.7 |
40-50 |
3860 |
| 41 |
53.4 |
50-60 |
3827 |
| 164 |
64.5 |
60-70 |
3881 |
| 24 |
74.7 |
70-80 |
3868 |
| 8 |
83.7 |
80-90 |
3868 |
| 平均 |
|
|
3860 |
若以每公斤体重的产热量进行比较,则小动物每公斤体重的产热量要比大动物高得多。若以每平方米体表面积的产热量进行比较,则不论机体的大小,各种动物每平方米每24小时的产热量很相近。在人体上测定的结果也表明了这一事实(表7-4)。因此,用每平方米体表面积标准来衡量能量代谢是比较合适的。
受试者体表面积的测定繁琐而不易进行,鉴于体表面积与身高、体重之间有一定的相关关系,因此,有人对一定的人群作过测定后,从身高、体重推算出体表面计算的经验公式。最基本的是Meeh的算式:
S=KW2/3
式中,S为体表面积,W为体重(kg),K为不同种属动物的常数。
计算人的体表面积在DuBois的身长体重算式:
S=W0.425×H0.725×K
式中,S为体表面积(m2),W为体重(kg),H为身长。K为不同人种的常数。我国人的体表面积可根据下列Stevenson算式来计算:
体表面积(m2)=0.0061×身长(cm)+0.0128×体重(kg)-0.1529
另外,体表面积还可根据图7-3直接求出。其用法是,将受试者的身高和体重在相应两条列线的两点连成一直线,引直线与中间的体表面积列线的交点就是该人的体表面积。有意义的事实是:肺活量、心输出量、主动脉和气管的横截面、肾小球滤过率等都与体表面积有一定的比例关系。]
通常采用前述简略法来测定和计算基础代谢率。下面举一实例供参考。
某受试者,男性,20岁。在基础状态下,1h的耗氧量为15L。非蛋白呼吸商定为0.82,氧热价为20.20,所以1h的产热量为:
20.20×15=303kJ
此人的体表面积为1.5m2,从而1h内每平方米体表面积拉热量,即基本代谢率为:303÷1.5=201.8kJ/m2・h
20岁男子的正常基础代谢率为157.8kJ/m2・h,所以此人超出正常值的数字为(201.8-157.8)=44.0,超出正常值的百分数为:
44.0×100/157.8%=27.8%
即+27.8%

图7-3 体表面积测算用图
实际测定结果表明,基础代谢率随性别、年龄等不同而有生理变化。当甚情况相同时,男子的基础代谢率平均比女子的高;幼年人比成年人的高;年龄越大,代谢率越低,但是,同一个体的基础代谢率,只在测定时的条件完全符合前述的要求,则有不同时日重复测定的结果基本上无差异。这就反映了正常人的基础代谢率是相当稳定的。
关于我国正常人基础代谢率的水平,男女各年龄组的平均值如表7-5所示。
一般来说,基础代谢率的实际数值现上述正常的平均值比较,相差±10%-15%之内,无论较高或较低,都不属病态。当相差之数超过20%时,才有可能是病理变化。在各种疾病中,甲状腺功能的改变总是伴有基础代谢率异常变化。甲状腺功能低下时,基础代谢率将比正常值低20%-40%;甲状腺功能亢进时的基础代谢率将比正常值高出25%-80%(图7-4)。因此,基础代谢率的测量是临床诊断甲状腺疾病的重要辅助方法。其它如肾上腺皮质和垂体的功能低下时,基础代谢率也要降低。
表7-5 我国人正常的基础代谢率平均值(kJ/m2・h)
| 年龄 |
11-15 |
16-17 |
18-19 |
20-30 |
31-40 |
41-50 |
51 以上 |
| 男性 |
195.5 |
193.4 |
166.2 |
157.8 |
158.7 |
154.1 |
149.1 |
| 女性 |
172.5 |
181.7 |
154.1 |
146.4 |
142.4 |
142.4 |
138.6 |
当人体发热时,基础代谢率将升高。一般说来,体温每升高1℃,基础代谢率可升高13%。其它如糖尿病、红细胞增多症、白血病以及伴有呼吸困难的心脏病等,也伴有基础代谢率升高。当机体处于病理性饥饿时,基础代谢率将降低。其他如阿狄森病、肾病综合症以及垂体肥胖症也常伴有基础代谢率降低。
图7-4 甲状腺疾病患者的基础代谢与正常人基础代谢的比较
纵坐标表示受检人数;横坐标的0代表正常平均值,
-10表示比正常人平均值低10%,10表示高10%