|
第三节 胃 内 消 化
胃是消化道中最膨大的部分。成人的容量一般为1-2L,因而具有暂时贮存食物的功能。食物入胃后,还受到胃液的化学性消化和胃壁肌肉运动的机械性消化。
一、胃 的 分 泌
胃粘膜是一个复杂的分泌器官,含有三咱管状外分泌腺和多种内分泌细胞。
胃的外分泌腺有:①贲门腺分布在胃与食管连接处的宽约1-4cm的环状区内,为粘液腺,分泌粘液;②泌酸腺分布在占全胃粘膜约2/3的胃底和胃体部。泌酸腺由三种细胞组成:壁细胞、主细胞和粘液颈细胞,它们分别分泌盐酸、胃蛋白酶原和粘液;③幽门腺分布在幽门部,是分泌碱性粘液的腺体。胃液是由这三种腺体和胃粘膜上皮细胞的分泌物构成的。
胃粘膜内至少含有6种内分泌细胞,如分泌胃泌素的G细胞、分泌生长抑素的D细胞和分泌组胺的肥大细胞等。
(一)胃液的性质、成分和作用
纯净的胃液是一种无色而呈酸性反应的液体,Pho 0.9-1.5。正常人每日分泌的胃液量约为1.5-2.5L。胃液的成分包括无机物如盐酸、钠和钾的氯化物等,以及有机物如粘蛋白、消化酶等。与唾液相似,胃液的成分也随分泌的速率而变化,当分泌率增加时,氢离子浓度升高,钠离子浓度下降,但氯和钾的浓度几乎保持恒定(图6-10)。
1.盐酸 胃液中的盐酸也称胃酸,其含量通常以单位时间内分泌的盐酸mmol表示,称为盐酸排出量。正常人空腹时盐酸排出量(基础酸排出量)约为0-5mmol/h。在食物或药物(胃泌素或组胺)的刺激下,盐酸排出量可进一步增加。正常人的盐酸最大排出量可达20-25mmol/h。男性的酸分泌多于女性;盐酸的排出量反映胃的分泌能力,它主要取决于壁细胞的数量(图6-11),但也与壁细胞的功能状态有关。

图6-10 人胃液中电解质浓度与分泌率的关系胃液分泌是用组胺静脉注射引起的
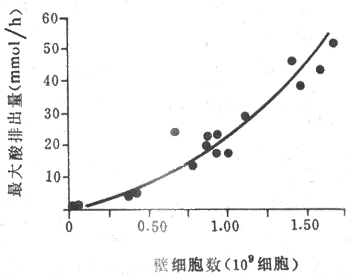
图6-11 胃酸最大排出量与壁细胞数目的关系
由17个人胃的切除部分计算出最大酸排出量和壁细胞数目的关系。
由图显示每100万个壁细胞可产酸约25mEp/h
胃液中H+的最大浓度可达150mmol/L,比血液中H+的浓度高三、四百万倍,因此,壁细胞分泌H+是逆着巨大的浓度梯度进行的,需要消耗大量的能量,能量来源于氧代谢。
泌酸所需的H+来自壁细胞浆内的水。水解离产生H+和OH-,任借存在于壁细胞上分泌小管膜上的H+、K+-ATP酶的作用,H+被主动地转运入小管腔内。
壁细胞分泌小管膜上的H+、K+-ATP酶又称质子泵(proton pump)或称酸泵。H+-K+交换是壁细胞质子泵区别于体内任何其它细胞上的质子泵的显著特征。H+、K+-ATP酶每催化一分子的ATP分解为ADP和磷酸所释放的能量,可驱动一个H+从壁细胞浆进入分泌小管腔和一个K+从小管腔进入细胞浆。H+的分泌必须在分泌小管内存在足够浓度的K+的条件下才能进行。
年来,选拔性干扰胃壁细胞的H+、K+-ATP酶的药物已被用来有效地抑制胃酸分泌,成为一代新型的抗溃疡药物。
已知壁细胞内含有丰富的碳酸酐酶,在它的催化下,由细胞代谢产生的CO2和由血浆中摄取的CO2可迅速地水合而形成H2CO3,H2CO3随即又解离为H+和HCO3。这样,在H+分泌后,留在细胞内的OH-便和由
H2CO3解离的H+结合而被中和,壁细胞内将不致因OH-的蓄积而使pH升高。由H2CO3产生的HCO3则在壁细胞的底侧膜,与CI-并换而进入血液。因此,餐后与大量胃酸分泌的同时,血和尿的pH往往升高而出现“餐后碱潮”。与HCO3交换而进入壁细胞内的CI-则通过分泌小管膜上特异性的CI-通道进入小管腔,与H+形成HCi (图6-12)。

图6-12 壁细胞分泌盐酸的一种假设
胃内的盐酸有许多作用,它可杀死随食物进入胃内的细菌,因而对维持胃和小肠内的无菌状态具有重要意义。盐酸还能激活胃蛋白酶原,使之转变为有活性的胃蛋白酶,盐酸并为胃蛋白酶作用提供了必要的酸性环境。盐酸进入小肠后,可以引起促胰液素的释放,从而促进胰液、胆汁和小肠液的分泌。盐酸所造成的酸性环境,还有助于小肠对铁和钙的吸收。但若盐酸分泌过多,也会对人体产生不利影响,。一般认为,过高的胃酸对胃和十二指肠粘膜有侵蚀作用,因而溃疡病发病的重要原因之一。
2.胃蛋白酶原 胃蛋白酶原是由主细胞合成的,并以不具有活性的酶原颗粒形式贮存在细胞内。当细胞内充满酶原颗粒时,它对新的酶原的全盛2产生负反馈作用。持续的刺激可使主细胞内的颗粒历释放而完全消失,便分泌仍继续进入,说明酶原也可以不经过颗粒的形式直接释放出来。
分泌入胃腔内的胃蛋白酶原在胃酸的作用下,从分子中分离出一个小分子的多肽,转变为具有活性的胃蛋白酶。已激活的胃蛋白酶对胃蛋白酶原也有激活作用。
胃蛋白酶能水解食物中的蛋白质,它主要作用于蛋白质及多肽分子中含苯丙氨酸或酪氨酸的肽键上,其主要分解产物是胨,产生多肽或氨基酸较少。胃蛋白酶只有在酸性较强的环境中才能发挥作用,其最知pH为2。随着pH的升高,胃蛋白酶的活性即降低,当pH升至6以上时,此酶即发生不可逆的变性。
3.粘液和碳酸氢盐 胃的粘液是由表面上皮细胞、泌酸腺的粘液颈细胞,贲门腺和幽门腺共同分泌的,其主要成分为糖蛋白。糖蛋白是由4个亚单位通过二硫键连接形成的(图6-13)。由于糖蛋白的结构特点,粘液具有较高的粘滞性和形成凝胶的特性。在正常人,粘液覆盖在胃粘膜的表面,形成一个厚约500μm的凝胶层,它具有润滑作用,可减少粗糙的食物对胃粘膜的机械性损伤。
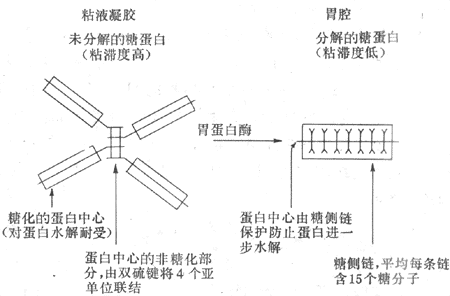
图6-13 胃粘液糖蛋白结构的示意图
胃内HCO3主要是由胃粘膜的非泌酸细胞分泌的,仅有少量的HCO3是从组织间液渗入胃内的。基础状态下,胃HCO3分泌的速率仅为H+分泌速率的5%。进食时其分泌速率的增加通常是与H+分泌速率的变化平行的。由于H+和HCO3在分泌速率和浓度上的巨大差距,分泌的HCO3对办内pH显然不会有多大影响。
长期以来人们一直在思索:胃粘膜处于高酸和胃蛋白酶的环境中,为什么不被消化?近年来“粘液-碳酸氢盐屏障”概念的提出,至少部分地回答了这个问题。这主要是因为。胃粘液的粘稠度约为水的30-260倍,H+和HCO3等离子在粘液层内的扩散速度明显减慢,因此,在胃腔同内的H+向粘液凝胶深层弥散过程中,它不断地与从粘液层下面的上皮细胞分泌并向表面扩散的HCO3遭遇,两种离子在粘液层内发生中和。用pH测量电极测得,在胃粘液层存在一个pH梯度,粘液层靠近胃腔面的一侧呈酸性,pH为7左右(图6-14)。因此,由沾液和碳酸氢盐共同构筑的粘液-碳酸氢盐屏障。能有效地阻挡H+的逆向弥散,保护了胃粘液免受H+的假侵蚀;粘液深层的中性pH环境还使胃蛋白酶丧失了分解蛋白质的作用。

图6-14 胃粘液-碳酸氢盐屏障模式图
正常情况下,胃粘液凝胶层临近胃腔一侧的糖蛋白容易受到胃蛋白酶的作用而水解为4个亚单位,这样,粘液便从凝胶状态变为溶胶状态而进入胃液。但一般来讲,水解的速度与粘膜上皮细胞分泌液的速度相等,这种粘液分泌与降解裼动态平衡,保持了粘液屏障功能的完整性和连续性。
4.内因子泌酸腺的壁细胞除分泌盐酸外,还分泌一种分子量在50000-60000之间的糖蛋白,称为内因子。内因子可与进入胃内的维生素B12结合而促进其吸收。
(二)胃液分泌的调节
胃液分泌受许多因素的影响,其中有的起兴奋性作用,有的则起抑制性作用。进食是胃液分泌的自然刺激物,它通过神经和体液因素调节胃液的分泌。
1.刺激胃酸分泌的内源性物质
(1)乙酰胆碱:大部分支配胃的副交感神经节后纤维末稍释放乙酰胆碱。乙酰胆碱直接作用于壁细胞膜上的胆碱能受体,引起盐酸分泌增加。乙酰胆碱的作用可被胆碱能受体阻断剂(如阿托品)阻断。
(2)胃泌素:胃泌素主要由胃窦粘膜内的G细胞分泌。十二指肠和空肠上段粘膜内也有少量G细胞。胃泌素释放后主要通过血液循环作用于壁细胞,刺激其分泌盐酸。
胃泌素以多种分子形式存在于体内,其主要的分子形式有两种:大胃泌素(G-34)和小胃泌素(G-17)。胃窦粘膜内的胃泌素主要是G-17,十二指肠粘膜中有G-17和G-34约各占一半。从生物效应来看,G-17刺激胃分泌的作用要比G-34强5-6倍,但G-34在体内被清除的速度很慢,它拦衰期约为50min,而G-17通常只有6min。
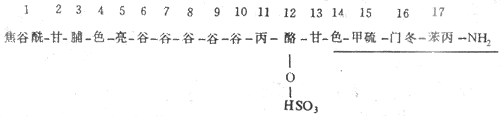
图6-15 胃泌素的氨基酸序列 划线的部分是其活性片段
人的小胃泌素的氨基酸序列如图6-15,其C端正的4个氨基酸是胃泌素的最小活性片段,因此,用人工合成的四肽或五肽胃泌素是具有天然胃泌全部作用的人工制品。
(三)组胺:胃的泌酸区粘膜内含有大量的组胺。产生组胺的细胞是存在于固有膜中的肥大细胞。正常情况下,胃粘膜恒定地释放少量组胺,通过局部弥散到达邻近的壁细胞,刺激其分泌。壁细胞上的组胺受体为Ⅱ型受体(H2受体),用甲氰咪呱(cimetidine)及其相类似的药物可以阻断组胺与壁细胞的结合,从而减少胃酸分泌。
以上三种内源性分泌物,一方面可通过各自壁细胞上的特异性受体,独立地发挥刺激胃酸分泌的作用(图6-16);另一方面,三者又相互影响,表现为当以上三个因素中的两个因素同时作用时,胃酸的分泌反应往往比这两个因素单独作用的总和要大,这种现象在生理学上称为加强作用(potentiation)。在整体内,促分泌物之间的相互加强作用是经常存在的,因此,用任何一咱促分泌物的阻断剂,如用甲氰咪呱时,它不仅抑制了壁细胞对组胺的反应,同时也会由于去除了组胺的作用的背景,使壁细胞对胃泌素和乙酰胆碱的反应也有所降低。
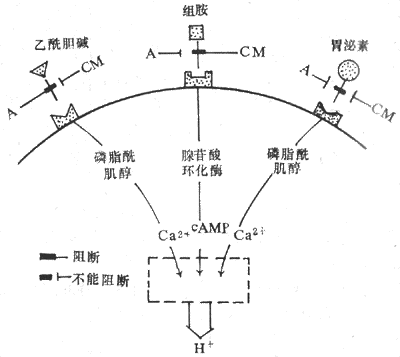
图6-16 三种刺激胃酸分泌的内源性物质的作用及相互关系
A:阿托品 CM:甲氰咪呱
2.消化期的胃液分泌 进食后胃液分泌的机制,一般按接受食物刺激的部位,分成三个时期来分析,即头期、胃期和肠期。但必须注意,三个时期的划分是人为的,只是为了便于叙述,实际上,这三个时期几乎是同时开始的、相互重叠的。
(1)头期胃液分泌:头期的胃液分泌是由进食动作引起的,因其传入冲动均来自头部感受器(眼、耳、口腔、咽、食管等)。因而称为头期。
头期胃液分泌的机制曾用慢性方法作了较详细的分析,即用事先旅行过食管切断术并具有胃瘘的狗进行假饲(sham feeding);当食物经过口腔进入食管后,随即从食管的切品流出体外,食物并未进入胃内,但却引起胃液分泌。进一步分析确定,由进食动作所引起的胃液分泌,包括条件反射性和非条例反射性两种分泌。前者是由和食物有关的形象、气味、声音等刺激了视、嗅、听等感受器而引起的;后者则当咀嚼和吞咽食物时,刺激了口腔和咽喉等外的化学和机械感受器而引起的。这些反射的传入途径和由进食引起的唾液分泌的传入途径相同,反射中枢包括延髓、下丘脑、边缘和大脑皮层等。迷走神经是这些反射共同的传出神经。当切断支配胃的迷走神经后,假饲就不再引起胃液分泌。
迷走神经兴奋后,除了通过其末稍释放乙酰胆碱,直接引起腺体细胞分泌外,迷走神经冲动还可引起胃窦粘膜内的G细胞释放胃泌素,后者经过血液循环刺激胃腺颁发(图6-17)。由此可见,头期的胃注分泌并不是纯神经反射性的,而是一种神经-体液性的调节。

图6-17 引起胃酸分泌的机制
引起胃泌素释放的迷走神经纤维被认为是非胆碱能的,因为阿托品不仅不能阻断,反而使假饲引起的胃泌素释放反应增加。目前对这一现象的解释是,迷走神经中既存在兴奋胃泌素释放的纤维,也存在抑制胃泌素释放的纤维,前者的中介物可能是一种肽类物质,而抑制性纤维则是通过乙酰胆碱能起作用的。阿托品由于阻断了抑制性纤维的作用,因而使胃泌素的释放有所增加。
头期胃液分泌的量和酸度都很高,而胃蛋白酶的含量则尤其高。在人体观察的资料表明,头期胃液分泌的大小与食欲有很大的关系。
(2)胃期胃液分泌:食物入胃后,对胃产生机械性和化学性刺激,继续引起胃液分泌,其主要途径为:①扩张刺激胃底、胃体部的感受器,通过迷走、迷走神经经长反射和壁内神经丛的短反射,引起胃腺分泌;②扩张刺激胃幽门部,通过壁内神经丛,作用于G细胞,引起胃泌素的释放;③食物的化学成分直接作用于G细胞,引起胃泌素的释放。
刺激G细胞释放胃泌素的主要食物化学成分是蛋白质的消化产物,其中包括肽类和氨基酸。G细胞为开放型胃肠内分泌细胞,顶端有绒毛样突起伸入胃腔,可以直接感受胃腔内化学物质的作用。用放射免疫方法测定血浆中胃泌素浓度,正常人空腹时约为30-120pg/ml,在进食蛋白质食物后,血浆胃泌素可升高到50-200pg/ml,在食后2-3小时逐渐恢复至进食前水平。糖类和脂肪类食物不是胃泌素释放的强刺激物。
胃酸分泌的胃液酸度也很高,但胃蛋白酶含量却比头期分泌的胃液为弱。
(3)肠期胃液分泌:将食糜、内的提取液、蛋白胨液由瘘管直接注入十二指肠内,也可引起胃液分泌的轻度增加,说明当食物离开胃进入小肠后,还有继续刺激胃液分泌的作用。机械扩张游离的空肠袢,胃液分泌也增加。
在切断支配胃的外来神经后,食物对小肠的作用仍可引起胃液分泌,提示肠期胃液分泌的机制中,神经反射的作用不大,它主要通过体液调节机制,即当食物与小肠粘膜接触时,有一种或几种激素从小肠粘膜释放出来,通过血液循环作用于胃。已知人的十二指肠粘膜中含有较多的胃泌素;用放射免疫方法测得,在切除了胃窦的病人,进食后血浆胃泌素的浓度仍有升高,说明进食后可引起十二指肠释放胃泌素,它可能是肠期胃泌分泌的体液因素之一。有人认为,在食糜作用下,小肠粘膜还可能释放肠泌酸素的激素,刺激胃酸分泌。此外,由小肠吸收的氨基酸也可能参与肠期的胃液分泌,因为静脉注射混合氨基酸也可引起胃酸分泌。
肠期胃液分泌的量不大,大约占进食后胃液分泌总量的1/10,这可能与食物在小肠内同时还产生许多对胃液起抑制性作用的调节(见后文)有关。
3.胃液分泌的抑制性调节上已述在进食过程中兴奋胃液分泌的机制,而正常消化期的胃液分泌还受到各种抑制性因素的调节,实际表现的胃液分泌正是兴奋和抑制性因素共同作用的结果。在消化期同内,抑制胃液分泌的因素除精神、情绪因素外,主要在盐酸、脂肪和高张溶液三种。
盐酸是胃腺活动的产物,但它对胃腺的活动又具有抑制性作用,因此是胃酸分泌的一种负反馈的调节机制。
当胃窦的pH降到1.2-1.5时,便可能对胃液分泌的产生抑制作用。这种抑制作用的机制可能是盐酸直接抑制了胃窦粘膜中的G细胞,减少胃泌素释放的结果。恶性贫血的病人胃酸分泌很低,他们血浆中胃泌素的浓度却比正常人高20-30倍,如向这种病人胃内注以盐酸,使胃内酸化,血浆胃泌素的浓度即下降,这进一步说明,胃内容物的酸度对胃泌素的释放,以及进而影响胃液分泌具有重要作用。
近年来一些实验资料提示,盐酸在胃内还可能通过引起胃粘膜释放一种抑制性因素――生长抑素,转而抑制胃泌素和胃液的分泌。
当十二接指肠内的pH降到2.5以下时,对胃酸分泌也有抑制作用,但其作用机制目前尚未完全阐明。已知酸作用于小肠粘膜可引起促胰液素释放,后者对胃泌素引起的酸分泌具有明显的抑制作用,因此,促胰液素很可能是十二指肠酸化抑制胃分泌的一种抑制物。此外,十二指肠球部在盐酸刺激下,也可能释放出一种抑制胃分泌的肽类激素――球抑胃素,但球抑胃素结构尚未最后确定。
脂肪是抑制胃液分泌的一个重要因素。脂肪及其消化产物抑制胃分泌的作用发生在脂肪进入十二指肠后,而不是在胃中。早在30年代,我国生理学家林可胜就发现,从小肠粘膜中可提取出一种物质,当由静脉注射后,保使胃液分泌的量、酸度和消化力减低,并抑制胃运动。这个物质被认为是脂肪在小肠内抑制胃分泌的体液因素,而可能是几种具有此种作用的激素的总称。小肠粘膜中存在的抑胃肽、神经降压素等多种激素,都具有类似肠抑素的特性。
十二指肠内高张溶液对胃分泌的抑制作用可能通过两种途径来实现,即汽船活小肠内渗透压感受器,通过肠-胃反射引起胃酸分泌的抑制,以及通过刺激小肠粘膜释放一种或几种抑制性激素而抑制胃液分泌。后一机制尚未阐明。
在胃的粘膜和肌层中,存在大量的前列腺素(详见内分泌章)。迷走神经兴奋和胃泌素都可引起前列腺素释放的增加。前列腺素对进食、组胺和胃泌素等引起的胃液分泌有明显的抑制作用。它可能是胃液分泌的负反馈抑制物。前列腺素还能减少胃粘膜血流,但它抑制胃分泌的作用并非继发于血流的改变。
二、胃 的 运 动
胃既有贮存食物的功能,又具有泵的功能。胃低和胃体的前部(也称头区)运动较弱,其主要功能是贮存食物;胃体的远端和胃窦(也称尾区)则有较明显的运动,其要主功能是磨粹食物、使食物与胃液充分混合,以形成食糜,以及逐步地将食糜排至十二指肠。
(一)胃的容受性舒张
当咀嚼和吞咽时,食物对回、食管等外感受器的刺激,可通过迷走神经反射性地引起胃底和胃体贴骨肉的舒张。胃壁肌肉的这种活动,被称为胃的容受性舒张。容受性舒张使胃腔容量由空腹时的50ml,增加到进食后的1.5L,它适应于大量食物的涌入,而胃内压力变化并不大,从而使胃更好地完成容受和贮存食物的功能,其生理意义是显然的。
胃的容受性舒张是通过迷走神经的传入和传出通路反射地实现的,切断人和动物的双侧迷走神经,容受性舒张即不再出现。在这个反射中,迷走神经和传也通路是抑制性纤维,其末稍释放的递质既非乙酰胆碱,也非去甲肾上腺素,而可能是某种肽类物质。
(二)胃的蠕动
食物进入胃后约5分钟,蠕动即开始。蠕动是从胃的中部开始,有节律地向幽门方向进行。在人,胃蠕动波的频率约每分钟3次,并需1分钟左右到达幽门。因此,通常是一波未平,一波又起。
蠕动波在初起时比较小,在向幽站传播过程中,波的深度和速度都逐步增加,当接近幽站时,明显加强,可将一部分食糜(约1-2ml)排入十二指肠,因此有幽站泵之称。并不是每一个蠕动波都到达幽门,有些蠕动波到胃窦后即行消失。一旦收缩波超越胃内容物,并到达胃窦终末时,由于胃窦终末部的有力收缩,胃内容物部分将被反向地推回到近侧胃窦和胃体部(图6-18)。食糜的这种后退,非常有利于食物和消化液的混合,还可机械地磨碎块状固体食物。总之,蠕动主要的生理意义是:一方面使食物与胃液充分混合,以利于胃液发挥消化作用;另一方面,则可搅拌和粉碎食物,并推进胃内容物通过幽门向十二指肠称行。
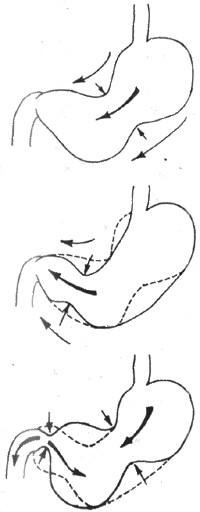
图6-18 胃的蠕动
胃的蠕动是受胃不滑肌的基本电节律控制的。胃的基本电节律起源于胃大弯上部,沿纵行肌向幽门方向传播,每分钟约3次。胃肌的收缩通常出现在基本电节律波后6-9s,动作电位后1-2s。神经和体液因素可通过影响胃的基本电节律和动作电位而影响胃的蠕动;迷走神经冲动、胃泌素和胃动素(是近年来从小肠粘膜中分离出来的一种胃肠激素)可使胃的基本电节律和动作电位出现的频率增加,使胃的收缩频率和强度增加;交感神经兴奋、促胰液素和抑胃肽则作用相反。
(三)胃的排空及其控制
食物由胃排入十二指肠的过程称为胃的排空。一般在食物入胃后5分钟即有部分食糜被排入十二指肠。不同食物的排空速度不同,这和食物的物理性状和化学组成都有关系。稀的、流体食物比稠的或固体食物排空快;切碎的、颗粒小的食物比大块的食物排空快;等涌液体比非等渗液体快。在三种主要食物中,糖类的排空时间较蛋白质为短,脂肪类食物排空最慢。对于混合食物,由胃完全排空通常需要4-6小时。
胃的排空率受来处胃和来自十二指肠二方面因素的控制:
1.胃内因素促进排空
(1)胃内食物量对排空率的影响:胃的内容物作为扩张胃的机械刺激,通过壁内神经反射或迷走-迷走神经反射,引起胃运动的加强。一般,食物由胃排空的速率和留在胃内作物量的平方根成正比。
(2)胃泌素对胃排空的影响:扩张刺激以及食物的某些成分,主要是蛋白质消化产物,可引起胃窦粘膜释放胃泌素。胃泌素除了胃酸分泌外,对胃的运动也有中等程度的刺激作用,它提高幽站泵的活动,便使幽站舒张,因而对胃拓空有重要的促进作用。
2.十二指肠因素抑制排空
(1)肠-胃反射对胃运动的抑制:在十二指肠壁上存在多种感受器,酸、脂肪、渗透压及机械扩张,都可刺激这些感受器,反射性地抑制胃运动,引起胃排空减慢。这个反射称为肠-胃反射,其传出冲动可通过迷走神经、壁内神经,甚至还可能通过交感神经等几条途径传到胃。肠-胃反射对酸的刺激特别敏感,当pH降到3.5-4.0时,反射即可引起,它抑制幽门泵的活动,从而阻止酸性食糜进入十二指肠。
(2)十二指肠产生的激素对胃排空的抑制:当过量的食糜,特别是酸或脂肪由胃进入十二指肠后,可引起不小粘膜释放几种不同的激素,抑制胃的运动,延缓胃的排空。促胰液素、抑胃肽等都具有这种作用,统称为肠抑胃素。
上述在十二指肠内具有抑制胃运动的各项因素产东是经常存在的,随着盐酸在肠内被中和,食物消化产物的被吸收,它们对胃的抑制性影响便渐渐消失,胃运动又逐渐增强,因而又推送另一部分食糜进入十二指肠。如此重复,使胃排内容物的排空较好地适应十二指肠内消化入吸收速度。
(四)呕吐
呕吐是将胃及肠内容物从口腔强力驱出的动作。机械的和化学的刺激作用于知根、咽部、胃、大小肠、总胆管、泌尿生殖器官等处的感受器,都可以引起呕吐。视觉和内耳庭的位置感觉发生改变时,也可引起呕吐。
呕吐前常出现恶习、流涎、呼吸急迫和心跳快而不规则等自主神经兴奋的症状。呕吐开始时,先是深吸气,声站紧闭,随着胃和食管下湍舒张,膈肌和腹肌猛烈地收缩,压挤胃的内容物通过食管而进入口腔。呕吐时,十二指肠和空肠上段也变得强烈起来,蠕动增快,并可转为痉挛。由于胃舒张而十二指肠收缩,平时的压力差转倒转,使十二指肠内容物倒流入胃,因此,呕吐物中常混有胆汁和小肠液。
在呕吐动作中,所有的这些活动都是反射性的。传入冲动的是由迷走神经和交感神经的感觉纤维、舌咽神经有及其它神经传入至延髓内的呕吐中枢。由中枢发出的冲动则沿迷走神经、交感神经、膈神经和脊神经等传到胃、小肠、膈肌和腹壁肌等处。呕吐中枢的位置在延髓外侧网状结构的背外侧缘。颅内压增高(脑水肿、肿瘤等情况)可直接刺激该中枢而引起呕吐。呕吐中枢在结构上和功能上与呼吸中枢、心血管中枢均有密切联系,它以协调这些邻近中枢的活动,从而在呕吐时产生复杂的反应。
在延髓呕吐中枢 的附近存在一个特殊的化学感受野,某些中枢性催吐药如阿朴吗啡,实际上是刺激了这个化学感受野,通过它再兴奋呕吐中枢的。
呕吐是一种具有保持意义的防御反射,它可把胃内有害的物质排出。但长期剧烈的呕吐会影响进食和正常消化活动,并且使大量的消化液丢失,造成体内水电解质和酸碱平衡的紊乱。
|