第六章 消化和吸收
第一节 概述
人的消化器官由长约8-10m的消化道及与其相连的许多大、小消化腺组成。消化器官的主要生理功能是对食物进行消化和吸收,从而为机体新陈代谢提供了必不可少的物质和能量来源。
消化是食物在消化道内被分解为小分子的过程。消化的方式有两种。一种是通过消化道肌肉的舒缩活动,将食物磨粹,并使之与消化液充分混合,以及将食物不断地向消化道的远端推送;这种方式称机械消化。另一种消化方式是通过消化腺分泌的消化液完成的。消化液中含在各种消化酶,能分别分解蛋白质、脂肪和糖类等物质,使之成为小分子物质(表6-1);这种消化方式称化学性消化。正常情况下,这两种方式的消化作用是同时进行,互相配合的。食物经过消化后,透过消化道的粘膜,进入血液和淋巴循环的过程,称为吸收。消化和吸收是两个相辅相成、紧密联系的过程。不能被消化和吸收的食物残渣,最后以粪的形式排出体外。
表6-1 消化液的成分及其作用
| 消化液 |
分泌量(L/d) |
PH |
主要成分 |
酶的底物 |
酶的水解产物 |
| 唾液 |
1.0-1.5 |
6.6-7.1 |
沾液 |
|
|
| |
|
|
α-淀粉酶 |
淀粉 |
麦芽糖 |
| 胃液 |
1.5-2.5 |
0.9-1.5 |
粘液、盐酸 |
|
|
| |
|
|
胃蛋白酶(原) |
蛋白质 |
、胨、多肽 |
| |
|
|
内因子 |
|
|
| 胰液 |
1.0-2.0 |
7.8-8.4 |
HCO3 |
|
|
| |
|
|
胰蛋白酶(原) |
蛋白质 |
氨基酸、寡肽 |
| |
|
|
糜蛋白酶(原) |
|
|
| |
|
|
羧基肽酶(原) |
肽 |
氨基酸 |
| |
|
|
核糖核酸酶 |
RNA |
单核苷酸 |
| |
|
|
脱氧核糖核酸酶 |
DNA |
|
| |
|
|
α-淀粉酶 |
淀粉 |
麦芽糖、寡糖 |
| |
|
|
胰脂肪酶 |
甘油三酯 |
脂肪酸、甘油、甘油一酯 |
| |
|
|
胆固醇酯酶 |
胆固醇酯 |
脂肪酸、胆固醇 |
| |
|
|
磷脂酶 |
磷脂 |
脂肪酸、溶血磷脂 |
| 胆汁 |
0.8-1.0 |
6.8-7.4 |
胆盐 |
|
|
| |
|
|
胆固醇 |
|
|
| |
|
|
胆色素 |
|
|
| 小肠液 |
1.0-3.0 |
7.6 |
粘液 |
|
|
| |
|
|
肠激酶 |
胰蛋白酶原 |
胰蛋白酶 |
| 大肠液 |
0.5 |
8.3 |
粘液 |
|
|
| |
|
|
HCO3 |
|
|
一、消化道平滑肌的特性
在整个消化道中,除口、咽、食管上端和肛门外括约肌是骨骼肌外,其余部分是都是由平滑肌组成的。消化道通过这些肌肉的舒缩活动,完成对食物的机械性消化,并推动食物的前进;消化道的运动对于食物的化学性消化和吸收,也有促进作用。
(一)消化道平滑肌的一般特性
消化道平滑肌具有肌组织的共同特性,如兴奋、自律性、传导性和收缩性,但这些特性的表现均有其自己的特点。
1.消化道平滑肌的兴奋较骨骼肌为低。收缩的潜伏期、收缩期和舒张期所占的时间比骨骼肌的长得多,而且变异很大。
2.消化道平滑肌在离体后,置于适宜的环境内,仍能进行良好的节律性运动,但其收缩很缓慢,节律性远不如心肌规则。
3.消化道平滑肌经常保持在一种微弱的持续收缩状态,即具有一定的紧张性。消化道各部分,如胃、肠等之所以能保持一定的形状和位置,同平滑肌的紧张性在重要的关系;紧张性还使消化道的管腔内经常保持着一定的基础压力;平滑肌的各种收缩活动也就是在紧张性基础上发生的。
4.消化道平滑肌能适应实际的需要而作很很大的伸展。作为中空的容纳器官来说,这一特性具有重要生理意义。它的消化道有可能容纳好几倍于自己原初体积的食物。
5.消化道平滑肌对电刺激较不敏感,但对于牵张、温度和化学刺激则特别敏感,轻微的刺激常可引起强烈的收缩。消化道平滑肌的这一特性是与它所处的生理环境分不开的,消化道内容物对平滑肌的牵张、温度和化学刺激是引起内容物推进或排空的自然刺激因素。
(二)消化道平滑肌的电生理特性
消化道平滑肌电活动的形式要比骨骼肌复杂得多,其电生理变化大致可分为三种,即静息膜电位、慢波电位和动作电位。
1.静息膜电位消化道平滑肌的静息膜电位很不稳定,波动较大,其实测值为-60―-50Mv,静息电位主要由K+的平衡电位形成,但Na+、CI-、Ca2+以及生电性钠泵活动也参与了静息膜电位的产生。
2.慢波电位消化道的平滑肌细胞可产生节律性的自发性去极化;以静息膜电位为基础的这种周期性波动,由于其发生频率较慢而被称为慢波电位,又称基本电节律(basal electric rhythm,BER)。消化道不部位的慢波频率不同,在人类,胃的慢波频率为3次/min,十二指肠为12次/min,回肠末端为8-9次/min。慢波的波幅约为10-15mV,持续时间由数秒至十几秒。
用细胞内微电极记录时,慢波多表现为单向波,包括初期的快速去极化和缓慢的复极化平台。关于慢波产生的离子基础尚未完全清楚。目前认为,它的产生可能与细胞膜上生电性钠泵的活动具有波动性有关,当钠泵的活动暂时受抑制时,膜便发生去极化;当钠泵活动恢复时,膜的极化加强,膜电位便又回到原来的水平。实验证明,用抑制钠泵的药物哇巴因后,胃肠平滑肌的慢波电位消失。
在通常情况下,慢波起源于消化道的纵行肌,以电紧张形式扩布到环行肌。由于切断支配胃肠的神经,或用药物阻断神经冲动后,慢波电位仍然存在,表明它的产生可能是肌源性的。慢波本身不引起肌肉收缩,便它可以反映平滑肌兴奋性的周期变化。慢波可使静息膜电位接近于产生动作电位的阈电位,一旦达到阈电位,膜上的电压依从性离子通道便开放而产生动作电位。
3.动作电位平滑肌的动作电位与神经和骨骼肌的动作电位的区别在于:①锋电位上升慢,持续时间长;②平滑肌的动作电位不受钠通道阻断剂的影响,但可被Ca2+通道阻断剂所阻断,这表明它的产生主要依赖Ca2+的内流;③不滑肌动作电位的复极化与骨骼肌相同,都是通过K+的外流,所不同的是,不滑肌K+的外向电流与Ca2+的内向电流在时间过程上几乎相同,因此,锋电位的幅度低,而且大小不等。
由于平滑肌动作电位发生时Ca2内流的速度已足以引起平滑肌的收缩,因此,锋电位与收缩之间存在很好的相关性,每个慢波上所出现锋电位的数目,可作为收缩力大小的指标。
慢波、动作电位和肌肉收缩的关系可简要归纳为:平滑肌的收缩是继动作电位之后产生的,而动作电位则是在慢波去极化的基础上发生的。因此,慢波电位本身虽不能引起平滑肌的收缩,但却被认为是不滑肌的起步电位,是平滑肌收缩节律的控制波,它决定蠕动的方向、节律和速度(图6-1)。

图6-1 消化道不滑肌的电活动
下面的曲线为细胞内电极记录的基本电节律,
在第二和第三个波的支极化期,出现数目不同的动作电位;
上面的曲线为肌肉收缩,收缩波只出现在动作电位时,
动作电位数目越多,收缩的幅度也越大T:张力 IP:细胞内电位
二、消化腺的分泌功能
人每日由各种消化腺分泌的消化液总量达6-8L(表6-1)。消化液主要由有机物、离子和水组成。消化液的主要功能为:①稀释食物,使之与血浆的渗透压相等,以利于吸收;②改变消化腔内的pH,使之适应于消化酶活性的需要;③水解复杂的食物成分,使之便于吸收;④通过分泌粘液、抗体和大量液体,保护消化道粘膜,防止物理性和化学性的损伤。
分泌过程是由腺细胞主动活动的过程,它包括由血液内摄取原料、在细胞内合成分泌物,以及将分泌物由细胞内排出等一连串的复杂活动。对消化腺分泌细胞的刺激-分泌耦联的研究表明,腺细胞膜上往往存在着多种受体,不同的刺激物与相应的受体结合,可引起细胞内一系列的生化反应,最终导致分泌物的释放(图6-2)。
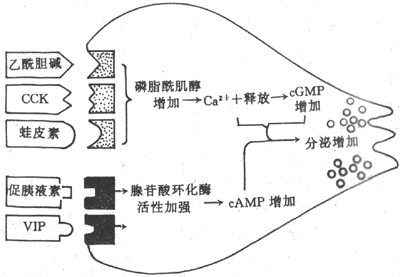
图6-2 胰腺分泌细胞内的两种介导机制
三、胃肠的神经支配及其作用
神经系统对胃肠功能的调节较为复杂,它通过植物性神经和胃肠的内在神经两个系统相互协调统一而完成的(图6-3)。
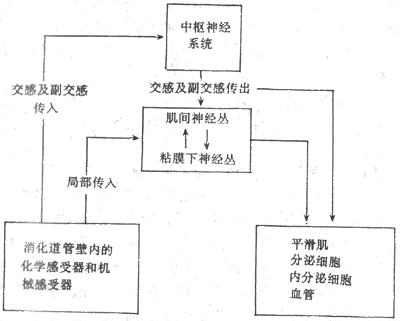
图6-3 消化系统的局部和中枢性反射通路
胃肠的内在神经是由存在于食管至肛门的管壁内的两种神经丛组成的。一种是位于胃肠壁粘膜下神经丛(Meissner神经丛);另一种是位于环行肌与纵行肌层之间的肌间神经丛(或称Auerbach神经丛)。内在神经丛包含无数的神经元和神经纤维,据估计,内在神经丛中约有108个神经元,包括感觉神经元、中间神经元和运动神经元。内在神经丛的神经纤维(包括进入消化管壁的交感和副交感纤维)则把胃肠壁的各种感受器及效应细胞与神经元互相连接,起着传递感觉信息、调节运动神经元的活动和启动、维持或抑制效应系统的作用。目前认为,消化管壁内的神经丛构成了一个完整的、相对独立的整合系统,在胃肠活动的调节中具有十分重要的作用。
支配胃肠的自主神经被称为外来神经,包括交感神经和副交感神经。交感神经从脊髓胸腰段侧角发出,经过腹腔神经节、肠系膜神经节或腥下神经节,更换神经元后,节后纤维分布到胃肠各部分,主要通过三种途径影响胃肠活动:①终止于内在神经元的肾上腺素能纤维;②分布于某些肌束的肾上腺素能纤维;③分布至血管平滑肌的肾上腺素血管纤维。由交感神经节后纤维释放至内在神经元表面的去甲肾上腺素,可抑制神经元的兴奋活动,从而抑制其向前传导的活动。这样,由交感神经发放的冲动,可抑制通过内在神经丛或迷走神经传递的反射。
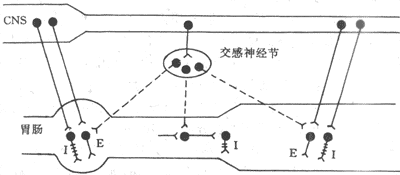
图6-4 胃肠胆碱能、肾上腺素能及肽能神经的支配及作用模式图
I:抑制 E:兴奋 CNS:中枢神经系统
――:胆碱能神经质 ----:肾上腺素能神经 +++肽能神经
副交感神经通过迷走神经和盆神经支配胃肠。到达胃肠的纤维都是节前纤维,它们终止于内在神经丛的神经元上。内在神经丛的多数副交感纤维是兴奋性胆碱能纤维,少数是抑制性纤维;而在这些抑制性纤维中,多数既不是胆碱能,也不是肾上腺素能纤维,它们的末梢释放的递质可能是肽类物质,因而被称为肽能神经(图6-4)。由肽能神经末梢释放的递质不是单一的肽,而可能是不同的肽,如血管活性肽(VIP)、P物质、脑啡肽和生长抑素等。目前认为,胃的容受性舒张、机械刺激引起的小肠充血等,均为神经兴奋释放VIP所致,VIP能神经的作用主要是舒张平滑肌、舒张血管和加强小肠、胰腺的分泌活动(图6-5)。

图6-5 血管活性肠肽(VIP)能神经支配的设想图
四、胃肠激素
在胃肠的粘膜层内, 不仅存在多种外分泌腺体,还含有十种内分泌细胞,这些细胞分泌的激素统称为胃肠激素(gastrointestinal hormone)。胃肠激素在化学结构上都是由氨基酸残基组成的肽类,分子量大多数在5000以内。
(一)胃肠内分泌细胞的形态及分布
用细胞免疫细胞化学的方法已经证明,从胃到大肠的粘膜层内,存在有40多种内分泌细胞,它们分散地分布在胃肠粘膜的跑非内分泌细胞之间。由于胃肠粘膜的面积巨大,胃肠内分泌细胞的总数很大有大,大大地超过了体内所有内分泌腺的总和。因此,消化道已不仅仅是人体内的消化器官,它也是体内最大最复杂的内分泌器官(表6-2)。
胃肠内分泌细胞在形成上有两个明显的特点,一是细胞内的分泌颗粒均分布在核和基底之间,故属于基底颗粒细胞。不同的内分泌细胞的分泌颗粒大小、形状和密度均不同。胃肠内分泌细胞的另一特点是,大部分细胞呈锥形,其顶端有绒毛突起,伸入胃肠腔内(图6-6),微绒毛可直接感受胃肠内食物成分和pH的一刺激而引起细胞的分泌活动。只有少数胃肠内分泌细胞无微绒毛,它们与胃肠腔无直接接触,它们的分泌可由神经兴奋或局部内环境的变化而引起,而与胃肠腔内的食物成分无关。这两种类型的细胞,前者被称为开放型细胞,后者为闭合型细胞。
表6-2 主要胃肠内分泌细胞的名称、分布和分泌产物
| 细胞名称 |
分泌产物 |
分布部位 |
| A细胞 |
胰高血糖素 |
胰岛 |
| B细胞 |
胰岛素 |
胰岛 |
| D细胞 |
生长 抑素 |
胰岛、胃、小肠、结肠 |
| G细胞 |
胃泌素 |
胃窦、十二指肠 |
| I细胞 |
胆囊收缩素 |
小肠上部 |
| K细胞 |
抑胃肽 |
小肠上部 |
| Mo细胞 |
胃动素 |
小肠 |
| N细胞 |
神经降压素 |
回肠 |
| PP细胞 |
胰多肽 |
胰岛、胰腺外分沁部分、胃、小肠、大肠 |
| S细胞 |
促胰液素 |
小肠上部 |

图6-6 胃窦粘膜内的G细胞(开放型细胞)示细胞顶端的绒毛
胃肠内分泌细胞在生物化学方面都具有摄取胺前体,进行脱羟而产生肽类或活性胺的能力。具有这种能力的细胞统称为APUD(amine precursor uptake and decarboxy-lation)细胞。除胃肠和胰腺的内分泌细胞外,神经系统、甲状腺、肾上腺髓质、垂体等组织中也含有APUD细胞。
(二)胃肠激素的作用
胃肠激素与神经系统一起,共同调节消化器官的运动、分泌和吸收功能。此外,胃肠激素对体内其它器官的活动也具有广泛的影响。其作用有三个主要方面。
1.调节消化腺的分泌和消化道的运动 这一作用的靶器官包括唾液腺、胃腺、胰腺、肠腺、肝细胞、食管-胃括约肌、胃肠平滑肌及胆囊等。三个主要胃肠激素的作用见表6-3及后文。
表6-3 三种胃肠对消化腺分泌和消化管运动地作用
| |
胃酸 |
胰HCO3 |
胰酶 |
肝胆汁 |
小肠液 |
食管-括约肌 |
胃运动 |
小肠运动 |
胆囊收缩 |
| 胃沁素 |
╂╂ |
╂ |
╂╂ |
╂ |
╂ |
╂ |
╂ |
╂ |
╂ |
| 促胰液素 |
― |
╂╂ |
╂ |
╂ |
╂ |
― |
― |
― |
╂ |
| 胆囊收缩素 |
╂ |
╂ |
╂╂ |
╂ |
╂ |
― |
╂ |
╂ |
╂╂ |
╂:兴奋 ╂╂:强兴奋 ―:抑制
2.调节其它激素的释放已经证明,食物消化时,从胃肠释放的抑胃肽(gastric inhibitory polypeptide,GIP)有很强的刺激胰岛素分泌的作用。因此,口服葡萄糖比静脉注射相同剂量的葡萄糖,能引起更多的胰岛素分泌。进餐时,不仅由于葡萄糖的吸收入血直接作用于胰岛B细胞,促进其分泌胰岛素,而且还可通过抑胃肽及早地把信息传递到胰岛,引起胰岛素较早的分泌;使血糖不至于升得过高而从尿中丢失,这对于有效地保持机体所获得的能源,具有重要的生理意义。
影响其它激素释放的胃肠激素还有:生长抑素、胰多肽、血管活性肽等,它们对生长激素、胰岛素、胰高血糖素、胃沁素等的释放均有调节作用。
3.营养作用 一些胃肠激素具有刺激消化道组织的代谢和促进生长的作用,称为营养作用(trophic action)。例如,胃泌素能刺激胃沁酸部和粘膜和十二指肠粘膜的蛋白质、RNA和DNA的合成。从而促进其生长。给动物长期注射五肽胃泌素(一种人工合成的胃泌素,含有胃泌素活性的最小片段―羧基端的5个氨基酸片段),可引起壁细胞增生。在临床上也观察到,切除胃窦的病人,血清胃泌素水平下降,同时可发生胃粘膜萎缩;相反,在患有胃泌素瘤的病人,血清胃泌素水平很高,这种病人多有胃粘膜增生、肥厚。此外,近年来还发现,小肠粘膜内I细胞释放的胆囊收缩素也具有重要的营养作用,它能引起胰腺内DNA、RAN和蛋白质的合成增加,促进胰腺外分泌组织的生长。
由胃肠内分泌细胞释放的激素主要是通过血液循环运送到靶细胞起作用的,这些出现在血液中的激素,可用放射免疫方法从血液中测定出来。但有一些胃肠激素释放后并不进入血液循环,而是通过细胞外注弥散至邻近的靶细胞,这种传递局部信息的方式也称为旁分泌(paracrine)。由胃窦部或胰岛内的D细胞释放的生长抑素,很可能是以这种方式发挥其对邻近的胃泌素细胞(G细胞)或胰B细胞的抑制性调节作用的。图6-7是胃肠激素这两种作用方式的模式图。

图6-7 胃肠激素作用的两种主要方式
A:经典的内分泌方式 B和C:旁分泌方式
(三)脑-肠肽的概念
近年来的研究证实,一些产生胃肠道的肽,不仅存在于胃肠道,也存在于中枢神经系统内;而原来认为只存在于中枢神经系统的神经肽,也在消化道中发现。这些双重分布的肽被统称为脑-肠肽(braingut peptide)。已知的脑-肠肽有胃泌素、胆囊收缩素、P物质、生长抑素、神经降压素等约20余种。这些肽类双重分布的生理意义已引起人们的重视,例如胆囊收缩素在外周对胰酶分泌和胆汁排放的调节作用及其在中枢对摄食的抑制作用,提示脑内及胃肠内的胆囊收缩素在消化和吸收中具有协调作用。