���Ľڡ�֬������л
����һ��֬����������ֽ�
����֬�������г���������������£��������ֽ�ΪCO2��H2O���ͷŴ������������֬�����ǻ�����Ҫ������Դ֮һ���κͼ����ǽ���֬�����������Ծ����֯��������Ҫ��������ʽ�Ǧ�-������
����(һ)֬����Ħ�-��������
�����˹��̿ɷ�Ϊ���ת�ƣ���-�����������Ρ�
����1.֬����Ļ
������������һ����֬����μӴ�лǰҲ��Ҫ�������ʽ��������֬����CoA����֬������ø��֬��CoA�ϳ�ø(acyl CoA synthetase)����
����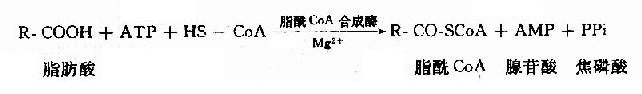
����������ɵ�֬��CoA������ǿ��������ˮ���������и��ܼ������ʻ��ã���ø����������ø����������˸����ײμӷ�Ӧ��
����֬��CoA�ϳ�ø�ֳ���ø���ֲ��ڰ����С�������Ĥ��������Ĥ�ϡ������е���ø��
�����ж���֬������������Ĥ�ϵ�ø�����֬���ᣬ����֬��CoA��Ȼ������������������������ϳɣ���������Ĥ�ϵ�ø��ij���֬��CoA����������������-������
����2.֬��CoA���������壺��֬�����-������øϵ������������У�������֬��CoA��������ͨ����������Ĥ��Ҫ������������ʾ���Ҫ����ת�ˣ���һ��������ⶾ��(carnitine)����3���ǣ�4����������������
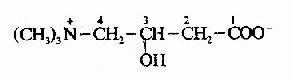
��������֬����CoA���ⶾ�Ӧ��������øA��֬���ⶾ�֬���������ⶾ���3���ǻ�ͨ�����������ӡ�
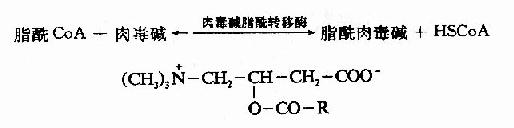
�������˷�Ӧ��øΪ�ⶾ��֬��ת��ø(carnitine acyl transferase)����������Ĥ������������д�ø��ϵͬ��ø���ֱ��Ϊ�ⶾ��֬��ת��øI���ⶾ��֬��ת��ø��ø��ʹ������֬��CoAת��Ϊ��øA��֬�����ⶾ����߽�����������Ĥ��λ����������Ĥ�ڲ��ø����ʹ֬�����ⶾ��ת�����ⶾ���֬��CoA���ⶾ�����·��������幦�ܣ�֬��CoA�������������ʣ���Ϊ֬�����-����øϵ�ĵ���(ͼ5��10)��
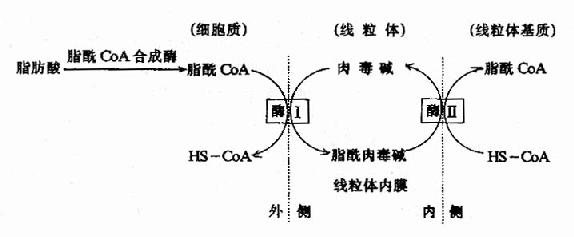
ͼ5-10���ⶾ�����֬����øAת��������ʾ��ͼ��
ø��λ����������Ĥ�����ⶾ��֬��ת��ø��
ø��λ����������Ĥ�ڲ���ⶾ��֬��ת��ø
��������֬��CoA������������ٶ��ܵ��ⶾ��֬��ת��ø���ø��ĵ��ڣ�ø���ܱ�����CoA���ƣ�ø�����ȵ������ơ�������CoA�Ǻϳ�֬�����ԭ�ϣ��ȵ���ͨ���յ�����CoA�Ȼ�ø�ĺϳ�ʹ������CoAŨ�����ӣ���������ø���Կ����ȵ��ض��ⶾ��֬��ת��ø���ø���м�ӻ�ֱ���������á��������ʳʱ�ȵ��ط��ڼ��٣��ⶾ��֬��ת��ø���ø��������ߣ�ת�Ƶij���֬��������������������ܡ�
����3.��-�����ķ�Ӧ���̣�֬��CoA������������н��������Ҫ�����IJ���Ӧ�������⡢��ˮ�����������⣬����һ��������CoA��һ��������̼���µ�֬��CoA��
������һ������(dehydrogenation)��Ӧ��֬��CoA����ø�������ΪFAD��֬��CoA�ڦ��ͦ�̼ԭ���ϸ���ȥһ����ԭ�����ɾ��з�ʽ˫���Ħ�����-ϩ֬������øA��

�����ڶ�����ˮ��hydration)��Ӧ��ϩ��CoAˮ��ø�������ɾ���L-���͵Ħ�-��֬��CoA��

�������������ⷴӦ���ڦ£���֬����CoA����ø����øΪNAD+�����£���-��֬����CoA�������ɦ�ͪ֬��CoA��

�������IJ���⣨thiolysis)��Ӧ�ɦ£�ͪ���ø�����£�ͪ����CoA�ڦ��ͦ�̼ԭ��֮�����������һ���Ӹ�øA��������CoA��һ��������̼ԭ�ӵ�֬��CoA��
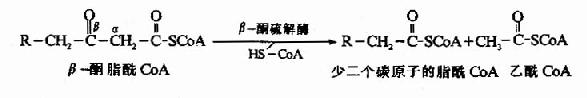
���������IJ���Ӧ��TCAѭ�����������ᾭ�Ӻ����ᡢƻ�������ɲ�������Ĺ������ƣ�ֻ�Ǧ�-�����ĵ��IJ���Ӧ����⣬�������������һ����Ӧ��������CoA�������������ᡣ
��������֬��CoA������һ��ѭ����̼����������̼ԭ�ӣ�����һ��������CoA������ظ������ѭ�����ͻ�����������CoA��
�������������Կ���֬����Ħ�-�������̾��������ص㡣����Ҫ��֬��������֬��CoA������һ�����ܹ��̡��С�����֬����������ֱ�����������壬������֬��CoA��Ҫ�ⶾ��ת�ˡ���-������Ӧ���������ڽ��У����û��������ĺ�ϸ����������֬���ṩ�ܡ���-������������FADH2��NADH+H+���ɣ���Щ��Ҫ�����������ݸ�������ˮ����Ҫ���μӣ�����CoA������Ҳ��Ҫ������ˣ���-�����Ǿ��������Ĺ��̡�
����֬�����-�������������̿�����ͼ(ͼ5��11)��ʾ��
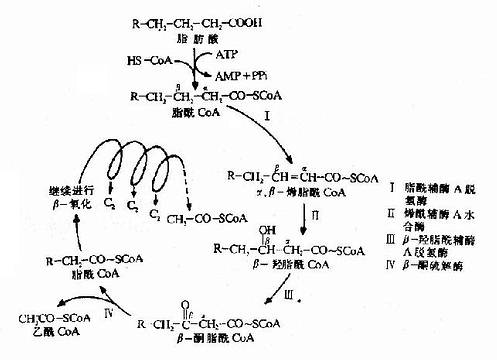
ͼ5-11��֬�����������Ӧ����
����(��)֬�����-��������������
����֬�����-����������֬����ֽ����Ҫ;����֬�����������Թ�Ӧ��������Ҫ�Ĵ�����������ʮ�˸�̼ԭ�ӵı���֬����Ӳ֬��Ϊ�������-�������ܷ�ӦΪ��
����CH3(CH2)15COSCoA+8NAD++*CoASH+8H2O������9CH3COSCoA+8FADH2+8NADH+8H+����
����8����FADH2�ṩ8��2=16����ATP��8����NADH+H+�ṩ8��3=24����ATP��9��������CoA��ȫ�����ṩ9��12=108������ATP�����һ�˷���Ӳ֬����ȫ��������CO2��H2O�����ṩ148�˷���ATP��Ӳ֬��Ļ��������2�˷���ATP������һ�˷���Ӳ֬����ȫ�����ɾ�����146�˷���ATP��һ�˷�����������ȫ����������38����ATP�����˷�������������̼ԭ������һ�˷���Ӳ֬����ͬ��ǰ�߿��ṩ114�˷���ATP�����߿��ṩ146�˷���ATP���ɼ���̼ԭ������ͬ�������֬�������ṩ�����������֬��������ʱ�ͷų���������Լ��40%Ϊ�������úϳɸ��ܻ��������60%���ȵ���ʽ�ͳ�����Ч��Ϊ40%��˵�������ܺ���Ч������֬�����������ṩ��������
����֬�����-����Ҳ��֬����ĸ�����̣���������Ҫ��֬�������ij��̲�ͬ��ͨ����-�����ɽ�����֬�������ɳ������˵�֬���ᣬ�������л���衣
����֬�����-�������������ɵ�����CoA��һ��ʮ����Ҫ���м仯�������CoA���ܽ���������ѭ�����������⣬����������Ҫ������ϳɵ�ԭ�ϣ���ͪ�塢���̴�����̴������
����(��)֬���������������ʽ
����1.����������������ں���ʳ�к�������������̼ԭ��֬���ᣬ������-��������������CoA�����һ���ӱ���CoA��ijЩ���������������ᡢ���������հ���ķֽ��л�������б���CoA���ɣ���֭�����ɹ��������������CoA������CoA�����Ȼ���Ӧ�ͷ��������ţ���ת������������CoA���ɽ�һ�������ֽ⣬Ҳ�ɾ����������������ǣ���Ӧ���̼���ͼ��

������������CoA��λø�ĸ�ø��5�䣭��������B12(5��dAB12)��ά����B12ȱ����5�䣭dAB12�����ϰ���Ӱ���λø���ԣ�ʹ��������CoA�ѻ��������һ�����������CoA��ȥ��øA�����ɼ�����������Ѫ�м������Ậ������(��������Ѫ֢)�����������ų�����(24Сʱ�ų�������4mgʱ��Ϊ����������֢)����һ�������������CoAŨ�����ߣ��ɲ���������֬��ϳɣ������쳣֬����(ʮ��̼��ʮ��̼��ʮ��̼֬����)���������������䡢����(�ٴ��ϳ�Ϊ�Ǽ��Ժϲ�����֢)��
����2.��-������֬����Ħ�-�������ڸ������н��У��ɼӵ���ø���ġ�������֬����Ħت�̼ԭ���ǻ����ɦ�-��֬���ᣬ�پ���ȩ֬�������ɦ�����-�����ᣬȻ���ڦ�-�˻��-�˻����������������-�������������������CoA��
����3.��-������֬�������������ɼӵ���ø������ø�����ɦ�-��֬�������һ��̼ԭ�ӵ�֬����Ĺ��̳�Ϊ֬����Ħ�-����������֬�����ɼӵ���ø�����ɿ���Ѫ�������Ҷ������������O2��Fe2+���������ɦ�-��֬���ᣬ��������֬����֬����Ҫ�ɷ֣���-��֬��������������Ⱦ���������̼ԭ��֬���ᡣ��-�����ϰ��߲�������ֲ����(phytanic acid,3��7��11��15-�ļ�ʮ������)��ţ�̺Ͷ���֬���о��д˳ɷ֣��������ڴ����ѻ�������Refsum�ϲ�����-������Ҫ������֯�ڷ����������-�����ϰ���������֢״��
����4.������֬����(unsaturated fatty acid)��������������Լ��1/2���ϵ�֬�����Dz�����֬���ᣬʳ����Ҳ���в�����֬���ᡣ��Щ������֬�����˫������˳ʽ�ģ����ǻ������-����ʱ������3��˳ϩ֬��CoA����ʱ��Ҫ˳��3����2�칹ø��ʹ������2����ϩ֬��CoA�Ա��һ����Ӧ��2����ϩ֬��CoA��ˮ������D����-��֬��CoA����Ҫ��-��֬��CoA�����칹ø����ʹ����D������ת���L�����ͣ��Ա��ٽ���������Ӧ(ֻ��L����-��֬��CoA������Ϊ��-��֬��CoA����ø�ĵ���)��
����������֬������ȫ��������CO2��H2Oʱ�ṩ��ATP������̼ͬԭ�����ı���֬���ᡣ
����(��)ͪ�������������
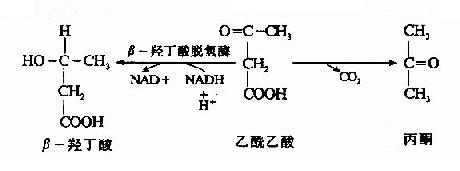
����ͪ��(acetone bodies)��֬�����ڸ�����������ֽ��л�����ɵ������м�����������������(acetoacetic acidԼռ30%)����-�Ƕ���(�ª�hydroxybutyric acidԼռ70%)�ͼ������ı�ͪ(acetone)��������ѪҺ��ͪ�庬������(ԼΪ0.8?.0mg/dl,0.2��2mM)��������������֬���������ܵ�����������ijЩ�������(��������ʳ)�����������(������)���ǵ���Դ�����������ϰ���֬��Ա��ǿ��֬����ͳ����������Ҫ�������ʡ������кϳ�ͪ���������������֯����ͪ�������������֮��ʧȥƽ�⣬Ѫ��Ũ�Ⱦͻ���ߣ�����ͪѪ֢(acetonemia)��ͪ��֢(acetonuria)����������ͦ�-�Ƕ��ᶼ���������ʣ����ͪ�������ڴ����ѻ������������ж���
����1.ͪ������ɹ��̣�
����ͪ�����ڸ�ϸ�������������ɵģ�������ԭ����֬�����-�������ɵ�����CoA�������Ƕ���������CoA�����ø��������ȥһ���Ӹ�øA��������������CoA������
������3���ǣ�3���������CoA(hydroxy methyl glutaryl��CoA,HMG��CoA)�ϳ�ø���£���������CoA����һ��������CoA��Ӧ������HMG��CoA�����ͷų�һ���Ӹ�ø����һ����Ӧ��ͪ�����ɵ����ٲ��衣
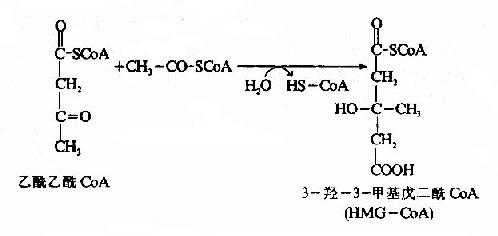
����HMG-CoA�ѽ�ø��HMG-CoA�����������������CoA�����߿�������ͪ��ĺϳɡ�
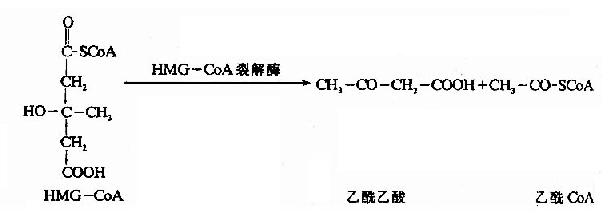
�����������еĦ£��Ƕ�������ø������������ԭ��NADH+H+�������壩�����ɦ�-�Ƕ��ᣬ�˻�ԭ�ٶȾ�������������[NADH+H+]/[NAD+]�ı�ֵ��������ջ��������������ɱ�ͪ��
��������ͪ�����ɹ���ʵ������һ��ѭ�����̣��ֳ�Ϊ����ѭ��(lynen cycle)��������������CoAͨ����ѭ������һ������������(��ͼ5��12)��

ͼ5-12��������ͪ�������
����ͪ�����ɺ�Ѹ������������Ĥ��ϸ��Ĥ����ѪҺ��ת����������֯���á�
����2.ͪ������ù���
�������������ļ�����������������CoAת��ø(succinyl��Coa thiophorase)����������CoA����ʱ����ø����������������������CoA����

�����ļ�����������л�����ø������ATP��øT����ʱ����ø��������������������CoA��

��������������ø�����ɵ���������CoA�����ø�����£��ֽ������������CoA������CoA��Ҫ����������ѭ�������ֽ⡣
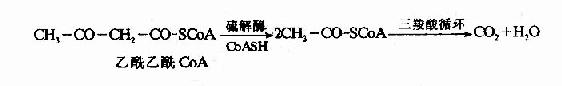
������ͪ�������ų��⣬��һ����ֱ�Ӵӷκ�������л�ϲ�ռ��Ҫ��λ��������֯������������ͦ�-�Ƕ���Ĺ��̿�����ͼ��ʾ(ͼ5��13)��

ͼ5-13��ͪ�����ù���
������ϸ����û��������CoAת��ø������������ø�����Ը�ϸ����������ͪ�塣
����������֯����ͪ������붯��Ѫ��ͪ��Ũ�ȳ����ȣ�����ͪ��Ũ�ȴ�70mg/dlʱ��������֯�����������ﵽ���͡���ͪ����Ϊ70mg/dl��Ѫ��ͪ��Ũ�ȳ�����ֵ��ͪ�徭��С����˹���������С�ܵ�����������������ͪ��֢������֯����ͪ���������Ѫ��ˮƽ�йأ�ֻ��Ѫ��ˮƽ����ʱ������ͪ�塣
����ͪ������ɺ����ù��̿�����ͼ��ʾ(ͼ5-14)��
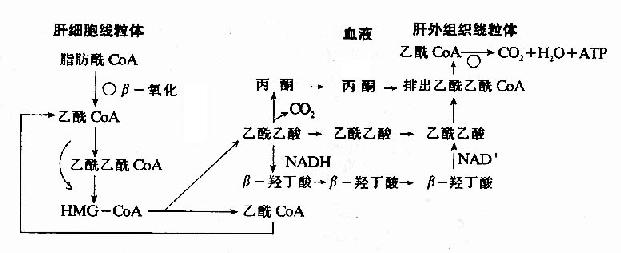
ͼ5-14��ͪ������ɺ�����
����3.ͪ�����ɵ�����
����(1)ͪ�������䣺����֬���ᴩ����������Ĥ��Ҫ�����ⶾ��ת�ˣ�֬������Ѫ��ת����Ҫ�������������֬����ף���ͪ��ͨ����������Ĥ�Լ���Ѫ��ת�˲�����Ҫ���塣
����(2)�����ã�֬����������-������ÿ��4����Ӧ��������һ��������CoA��������������ֻ��һ����Ӧ�Ϳ�����������������CoA����-�Ƕ��������ֻ�����������һ��������Ӧ����ˣ�����ͪ�忴����֬�����ڸ���ӹ����ɵİ��Ʒ��
����(3)��ʡ�����ǹ��Ժͺ�ϸ�����ã�������֯����ͪ������ɴ���������CoA����������CoA
�������Ʊ�ͪ������øϵ���ԣ������ǵ����á�ͬʱ����CoA���ܼ����ͪ���Ȼ�ø���ٽ���������������֯����ͪ���������ܣ��ͼ����˶������ǵ������Ա�֤����֯����ϸ���������ǵ���Ҫ������֯�������ó���֬���ᣬ���ڼ���ʱ������ͪ�幩�ܣ�����5?��ʱͪ�幩�ܿɶ��70%��
����(4)������֯����ͪ�壬�������Ƽ��⵰���ʵķֽ⣬��ֹ�����ʹ������ģ������û����в������
����(5)ͪ���������ೣ���ڼ����������ж�֢����������¡����Ǹ�֬��ʳҲ��ʹͪ���������ࡣ
��������֬����ĺϳ�
���������ڵ�֬�������Դ��ʳ�Ϊ��Դ��֬���ᣬ�����ڿ�ͨ������ӹ����������á�ͬʱ���廹���������Ǻ͵���ת��Ϊ֬�����Ϊ��Դ��֬���ᣬ���ڸ������������ɣ������������ϳ�֬�������Ҫ�����Ǹ���Ͳ���������������֬����֯�����ࡢС�������Ժϳ�֬���ᣬ�ϳ�֬�����ֱ��ԭ��������CoA������ATP��NADPH����������ʮ��̼����֬�ᣬ�����ӹ������������֬���ᣬ�ϳ���ϸ�����н��С�
����(һ)��֬�������
����֬����ĺϳ�����������CoA��ʼ�ϳɣ�������ʮ��̼�ı���֬���ἴ������(palmitoleic acid)��
����1.����CoA��ת��
��������CoA�����������ֽ����֬���ᡢͪ��͵��ֽ����ɣ���������CoA�ķ�Ӧ���������������У���֬����ĺϳɲ�λ�ǰ������������CoA������������ת������������������CoA��������ͨ��������Ĥ����Ҫͨ��һ����Ϊ�����᪲��ͪ��ѭ��(citrate pyruvate cycle)���������CoA�������嵽������ת�ơ��������������ڣ�����CoA��������ᾭ������ϳ�ø�����������������ᣬ������������Ĥ����Ӧ����Э�������Һ���ڰ�Һ�ڴ��ڵ��������ѽ�ø(citrate lyase)��ʹ�������ѽ��������CoA���������ᡣǰ������������֬���ᣬ���߿ɷ��������岹��ϳ�������ʱ�����ġ�����������Ҳ��������ͨ��������Ĥ���ʱ����Ⱦ�ƻ��������ø������ԭ��ƻ�����پ���������Ĥ�ϵ�����ת���������壬��������������ᡣҲ����ƻ����ø�����£������������ɱ�ͪ�ᣬͬʱ����NADPH�����ɡ���ͪ��ɾ���Ĥ���屻ת�����������ڣ���ʱ��ͪ������Ȼ�ת��Ϊ�������ᡣÿ���������ͪ��ѭ��һ�Σ���ʹһ��������CoA������������Һ��ͬʱ����������ATP����Ϊ�����ṩ��NADPH�Բ���ϳɷ�Ӧ����Ҫ(��ͼ5-15)��
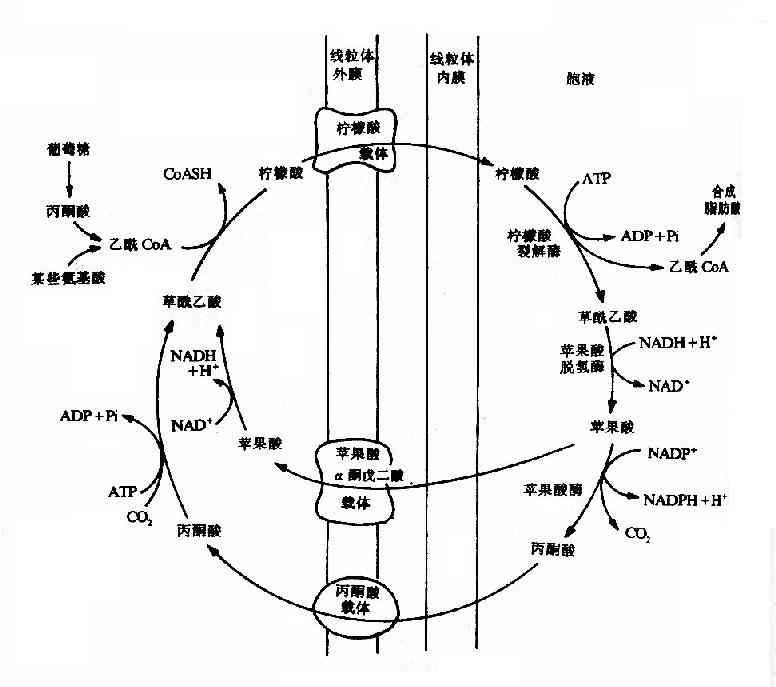
ͼ5-15��������-��ͪ��ѭ��
����2.������CoA������
��������CoA������CoA�Ȼ�ø(acetyl CoA carboxylase)��ת��ɱ�����CoA(��Ʊ����ᵥ��CoA)��Ӧ���£���������

��������CoA�Ȼ�ø�����ڰ�Һ�У��丨��Ϊ�����أ��ڷ�Ӧ��������Я����ת���Ȼ������á��÷�Ӧ�����������������������ص��Ȼ���Ӧ�������ͪ���Ȼ���Ϊ��������ķ�Ӧ�ȡ�
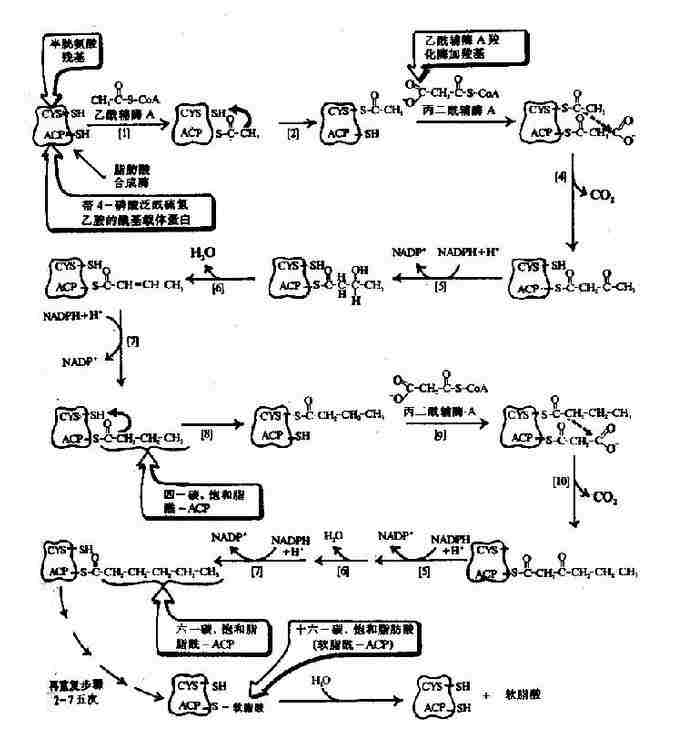
��ͼ5��16��ԭ������֬����ϳ�ø������������֬��(16��0)
����������CoA�Ȼ�ø���ķ�ӦΪ֬����ϳɹ����е����ٲ��衣��øΪһ��ø���ڱ乹ЧӦ���������£������Եĵ������л��ԵĶ����(��10?0���������״����)֮����Ի��䡣����������������ɴٽ�����ۺϳɶ���壬��ǿø���ԣ�������֬����ɼ��ٽ�ۣ��Ӷ����Ƹ�ø���ԡ�����CoA�Ȼ�ø����ͨ��������cAMP�����ữ��ȥ���ữ����������ø���ԡ���ø�����ữ�����ɥʧ��������Ѫ�����������������ܴٽ��������ữ���ã��Ӷ�����֬����ϳɣ����ȵ������ܴٽ�ø��ȥ���ữ���ã��ʿ���ǿ����CoA�Ȼ�ø���ԣ�����֬����ϳɡ�
����ͬʱ����CoA�Ȼ�øҲ���յ�ø�����ڸ��ǵ�֬��ʳ���յ���ø���ɣ��ٽ�֬����ϳɣ���֮����֬������ʳ�����ƴ�ø�ϳɣ�����֬��������ɡ�
����3.��֬�������
������֬��ĺϳ�ʵ������һ���ظ�ѭ���Ĺ��̣���1��������CoA��7���ӱ�����CoA��ת�ơ����ϡ����⡢��ˮ���ټ����ظ����̣�ÿһ��ʹ̼���ӳ�����̼����7���ظ����������ɺ�ʮ��̼����֬��(ͼ5-16)��

������ԭ������(��˾���)���˷�Ӧ��ø��һ����7�ֲ�ͬ���ܵ�ø��һ���������嵰��(acyl carrier protein��ACP)�ۺϳɵĸ����塣�����������˷�Ӧ��һ�ֺ���˫�ǻ���ø��ÿ���ǻ���7����ͬ�����ܵĽṹ����һ���൱��ACP�Ľṹ�����������һ�־��ж��ֹ��ܵ�ø��
����֬����ϳ�������ATP��NADPH+H+,NADPH��Ҫ��Դ�������Ƿֽ����������;�������⣬ƻ������������Ҳ�ɲ�������NADPH��
����֬����ϳɹ��̲��Ǧ�-����������̣����Ƿ�Ӧ����֯��ϸ����λ��ת�����壬�������壬����ø������������Ƽ�����������������Լ���Ӧ��������������ͬ(��5-6)��
����5-6��֬����ϳɺͷֽ�ıȽ�
| �� |
�ϳ� |
�ֽ� |
| ��Ӧ���Ծʱ�� |
������ʳ�� |
���� |
| �̼����� |
�ȵ���/�ȸ�Ѫ���ظ߱�ֵ |
�ȵ��أ��ȸ�Ѫ���صͱ�ֵ |
| ��Ҫ��֯��λ |
������ |
���⡢���� |
| ��ϸ����λ |
���� |
�������� |
| �������� |
�����ᣨ�����嵽������ |
�ⶾ������������壩 |
| ������������Ұ��Ļ������� |
�������嵰������CoA |
CoA |
| ������ԭ������ |
NADPH |
NAD+��FAD |
| ��̼����/���� |
������CoA���������� |
����CoA������ |
| �����
���Ƽ� |
������֬��øCoA����������CoA�Ȼ�ø�� |
������CoA�������ⶾ������ת��ø�� |
| ��Ӧ���� |
��֬�� |
������øA |
����(��)����֬���������
���������ڲ�������֬�ᣬ����̼�����̲��ȵ�����֬���ᣬҲ�и��ֲ�����֬���ᣬ��Ӫ������֬��������ʳ�﹩Ӧ�⣬����֬�����������֬����ϸ���ڼӹ�������ɡ�
����1.̼�����ӳ�������
����֬����̼�����������������о���-������ɣ�����һ�Φ�-����ѭ���Ϳ��Լ�������̼ԭ�ӡ�
����֬����̼�����ӳ����ڻ������������������о�֬�����ӳ�ø��ϵ����ɡ�
������������������֬���ӳ����Ա�����CoAΪ��̼��λ�Ĺ��壬��NADPH+H+���⣬�ྭ�������ȡ���ԭ�ȹ����ӳ�̼�������Һ��֬����ϳɹ��̻�����ͬ��������Ӧ��ø��ϵ��ͬ����֬������������ACPΪ���壬�����븨øA�����μӷ�Ӧ��������֯��һ���Ժϳ�Ӳ֬��(18C)Ϊ��������֯������ø���ʿ��ӳ���24̼��֬���ᣬ������֬���л��Ҫ��
�����������壬��֬�ᾭ������֬�����ӳ�ø��ϵ���ã�������CoA�������ӳ�̼�����������֬������������з�Ӧ���ƣ���ϩ֬��CoA��ԭø�ĸ�øΪNADPH+H+����������̲�ͬ��ͨ�����ַ�ʽһ����ӳ�֬����̼����24��26̼������Ӳ֬����ࡣ
����2.֬�����ѱ���
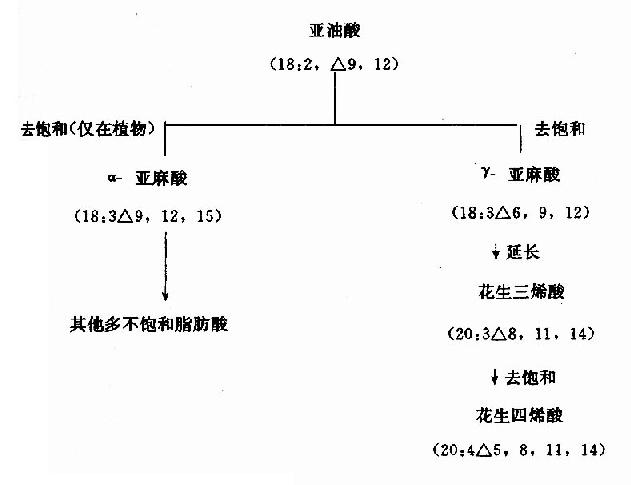
�����˺Ͷ�����֯���еIJ�����֬������ҪΪ������(16��1��9)������(18��1��9)��������(18��2��9��12)��������(18��3��9��12��15)��������ϩ��(20��4��5��8��11��14)�ȡ���������ͨ�ĵ�������֬���᪩������������������Ӧ��֬������ȥ����ø(acylCoAdesaturase)���������ɡ�����ø�����ڻ���������������Ϲ�������ø(�������л��)�����øֻ���ڡ�9�γ�˫������������C10��ĩ�˼�֮���γ�˫������������(linoleate)��������(linolenate)��������ϩ��(arachidonate)�����ڲ��ܺϳɻ�ϳɲ��㡣���������ǻ��岻��ȱ�ٵģ����Ա�����ʳ�﹩������ˣ���֮Ϊ����֬����(essential fatty acid)��ֲ����֯���п�����C��10��ĩ�˼����γ�˫��(����3�ͦ�6)��ȥ����ø���ܺϳ�����3�ֶ����֬���ᡣ��ʳ����������ڶ������ھ�̼���ӳ���ȥ���ͺ����ɻ�����ϩ�ᡣ
����(��)֬����ϳɵĵ���
��������CoA�Ȼ�ø���ķ�Ӧ��֬����ϳɵ����ٲ��裬�ܶ����ض���Ӱ���ø���ԣ��Ӷ�ʹ֬����ϳ��ٶȸı䡣֬����ϳɹ���������ø����֬����ϳ�ø���������ѽ�ø����ɱ����ڡ�
����1.��л��ĵ���
�����ڸ�֬��ʳ��������֬����Ա��ǿʱ��ϸ������֬��CoA���࣬�ɷ�����������CoA�Ȼ�ø���Ӷ���������֬����ϳɡ�����ʳ���࣬�Ǵ�л��ǿʱ��������������������ѭ���ṩ������CoA��NADPH���࣬��Щ�ϳ�֬�����ԭ�ϵ�����������֬����ĺϳɡ����⣬��������ǿ�Ľ����ʹϸ����ATP���࣬��������������������ø������������ἰ������ѻ�������������Ĥ����Ӧ����Э���£���������ת���Һ�����Ա�������CoA�Ȼ�ø��ͬʱ����Ҳ���ѽ��ͷ�����CoA������֬����ϳɵ�ԭ�ϣ�ʹ֬����ϳ����ӡ�
����2.���صĵ���
�����ȵ��ء��ȸ�Ѫ���ء��������ؼ������صȾ������֬����ϳɵĵ��ڡ�
�����ȵ������յ�����CoA�Ȼ�ø��֬����ϳ�ø���������ѽ�ø�ĺϳɣ��Ӷ��ٽ�֬����ĺϳɡ����⣬����ͨ���ٽ�����CoA�Ȼ�ø��ȥ���ữ��ʹø������ǿ��Ҳʹ֬����ϳɼ��١�
�����ȸ�Ѫ���صȿ�ͨ������cAMP����ʹ����CoA�Ȼ�ø���ữ�����ͻ��ԣ��������֬����ĺϳɡ����⣬�ȸ�Ѫ����Ҳ���Ƹ��������ϳɣ��Ӷ����ӳ���֬��CoA������CoA�Ȼ�ø�ķ������ƣ���ʹ֬����ϳɱ����ơ�
����(��)ǰ�����ء�Ѫ˨�ؼ�����ϩ
����ǰ������(prostaglandin��PG)��Ѫ˨��(thromboxane��TX)�Ͱ���ϩ(leukotrienes,LT)���ɻ�����ϩ������������������ϸ�������ɺ���Ϊ������Լ������е�ϸ����л���ӵ������ã���������֢��������Ӧ����Ѫ�������Ȳ��������йء�
��������Ĥ�ϵ�Ĥ��֬���л�����ϩ�ᣬ���ɱ���֬øA2ˮ�⣬�ͷŻ�����ϩ�ᡣ������ϩ�����ǰ�������ڹ�������ϳ�ø���£�����O2�ͻ�ԭ�������������������ͻ�����Ӧ������ǰ������H2��ǰ������H2�ɽ�һ������������ǰ�����ؼ�Ѫ˨��(��ͼ5?7)���ɵ���(cortisol)��������øA2���ԣ����ٻ�����ϩ������ɣ��Ӷ�����ǰ��ԭ�صĺϳɣ���˹ƥ��(aspirin)����̩��(phenylbutazone)����ǰ�������ڹ�������ϳ�ø����ʹǰ�����غ�Ѫ˨�����ɼ��١�

ͼ5-17��������ϩ������PG��TX��LT�ſ�
��������PG��TX��LT����������
����PG����ϸ���ں����ܵͣ���10?1pmol/L�������к�ǿ���������ԡ�
����(һ)PG
����PGE2���շ���֢���ٽ��ֲ�Ѫ�����ţ�ëϸѪ��ͨ�����ӣ�����졢�ס�ʹ���ȵ�֢״��PGE2��PGA2ʹ����ƽ�������ţ��н���Ѫѹ�����ã�PGE2��PGI2����θ����ڣ��ٽ�θ��ƽ�����䶯�����ݲ�����PGE2��PGE2�������ѹ���������Ҫ���ã�PGE2����ʹ�ѳ�ƽ�����������������ѡ��ӹ��ͷŵ�PGE2����ʹ�����ܽ⣬����ʱ�ӹ���Ĥ�ͳ���PGE2���������ӹ�������ǿ���ٽ����䡣
����(��)TX
����ѪС�������TXA2��PGE2�ٽ�ѪС��ۼ���Ѫ���������ٽ���Ѫ��Ѫ˨�γɣ���Ѫ����Ƥϸ���ͷŵ�PGI2���к�ǿ����Ѫ�ܼ���ѪС��ۼ���������Ѫ��Ѫ˨�γɣ���TXA2�����öԿ�������������˹��Ħ����ʳ����������ϩ���Ѫ��ʳ��������ںϳ�PGE3��PGI3��TXA3��������PGI3�����ƻ�����ϩ���Ĥ��֬�ͷţ��������PGI2��TXA2�ĺϳɡ�����PGI3�Ļ�����PGI2��ͬ����TXA3���TXA2���ö࣬��˰�˹��Ħ�˿�ѪС��ۼ�������Ѫ���ý�ǿ������Ϊ�����Dz����ļ���������Ҫԭ��֮һ��
����(��)LT
������֤ʵ������Ӧ������Ӧ����(SRS��A)��LTC4��TD4��LTE4�Ļ�����ʹ֧����ƽ�������������ý��鰷��PGF2ǿ100000�������û������־á����⣬LTG4���ܵ��ڰ�ϸ���Ĺ��ܣ��ٽ��������������ã��̼������ỷ��ø���շ���˰�ϸ���ѿ�����ʹ��ø�ͷ�ˮ��ø�����ٽ���֢��������Ӧ�ķ�չ����