ЕкЖўНкЁЁбЊжЌМАЦфДњаЛ
ЁЁЁЁбЊНЌжаКЌгаЕФжЌРрЭГГЦЮЊбЊѕЅЃЌАќРЈИЪгЭШ§ѕЅЁЂСзжЌЁЂЕЈЙЬДММАЦфѕЅКЭЗЧѕЅЛЏжЌЗО(nonЊВesterified fatty acid)ЃЌврГЦгЮРыжЌЗОЫс(free fatty acid,МђаДFFA)ЁЃбЊжЌдкжЌРрЕФдЫЪфКЭДњаЛЩЯЦ№зХживЊзїгУЁЃбЊжЌжЛеМЬхжиЕФ0.04%ЃЌЦфКЌСПЪмЕНвћЪГЁЂгЊбјЁЂМВВЁЕШвђЫиЕФгАЯьЃЌвђЖјЪЧСйДВЩЯСЫНтЛМепжЌРрДњаЛЧщПіЕФвЛИіживЊДАПкЁЃе§ГЃШЫбЊжЌКЌСПМћБэ5?ЁЃЫќУЧЪЧвджЌЕААзЕФаЮЪНДцдкВЂдЫЪфЕФЃЌжЌЕААзгЩжЌРргыдижЌЕААзНсКЯЖјаЮГЩЁЃжЌЕААзОпгаЮЂЭХНсЙЙЃЌЗЧМЋадЕФИЪгЭШ§ѕЅЁЂЕЈЙЬДМѕЅЕШЮЛгкКЫаФЃЌЭтжмЮЊЧзЫЎадЕФдижЌЕААзКЭЕЈЙЬДМСзжЌЕШЕФМЋадЛљвђЃЌетбљЪЙжЌЕААзОпгаНЯЧПЫЎШмадЃЌПЩдкбЊвКжадЫЪф(ЭМ5Ѓ2)ЁЃ
Бэ5ЃЃБЁЁе§ГЃГЩШЫПеИЙбЊжЌЕФжївЊГЩЗжКЭКЌСП
| жЌРрЮяжЪ |
КЌСПЃЈКСПЫЃЏ100КСЩ§бЊНЌЃЉ |
жЌРрЮяжЪ |
КЌСПЃЈКСПЫЃЏ100КСЩ§бЊНЌЃЉ |
| жЌРрзмжи |
400-700ЃЈ500ЃЉ |
ЕЈЙЬДМ |
105-260ЃЈ200ЃЉ |
| ИЪгЭШ§ѕЅ |
10-160ЃЈ100ЃЉ |
ѕЅаЭ |
90-260ЃЈ145ЃЉ |
| СзжЌ |
150-250ЃЈ200ЃЉ |
здгЩаЭ |
40-70ЃЈ55ЃЉ |
| СзжЌѕЃЕЈМю |
80-225ЃЈ110ЃЉ |
жЌЗОЫсзмСП |
110-485ЃЈ300ЃЉ |
| СзжЌѕЃввДМАЗ |
0-30ЃЈ10ЃЉ |
ЗЧѕЅЛЏаЭЛЏжЌЗОЫс |
5-20 |
| ЩёОСзжЌ |
10-50ЃЈ30ЃЉ |
ЁЁ |
ЁЁ |
ЁЁЁЁзЂЃКРЈЛЁФкЮЊЦНОљжЕЊЄ

ЭМ5-2ЁЁбЊНЌжЌЕААзЕФвЛАуНсЙЙ
ЁЁЁЁвЛЁЂбЊНЌжЌЕААзЕФЗжРр
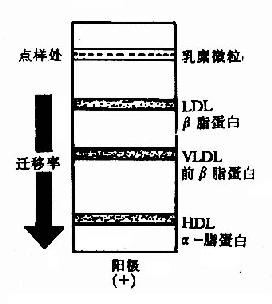
ЭМ5-3ЁЁбЊНЌжЌЕААзЕФЕчгОааЮЊ
ЁЁЁЁбЊвКжаЕФжЌЕААзВЛЪЧЕЅвЛЕФЗжзгаЮЪНЃЌЦфжЌРрКЭЕААзжЪЕФзщГЩгаКмДѓЕФВювьЃЌвђДЫбЊвКжаЕФжЌЕААзДцдкЖржжаЮЪНЁЃИљОнЫќУЧИїздЕФЬиадВЩгУВЛЭЌЕФЗжРрЗНЗЈЃЌПЩНЋЫќУЧНјааЖржжЗжРрЃЌвЛАуВЩгУЕчгОЗЈКЭГЌЫйРыаФЗЈНјаабЊНЌжЌЕААзЕФЗжРрЁЃ
ЁЁЁЁ(вЛ)ЕчгОЗжРрЗЈ
ЁЁЁЁБОЗЈИљОнВЛЭЌжЌЕААзЫљДјБэУцЕчКЩВЛЭЌЃЌдквЛЖЈЭтМгЕчГЁзїгУЯТЃЌЕчгОЧЈвЦТЪВЛЭЌЃЌПЩНЋбЊНЌжЌЕААзЗжЮЊЫФРрЁЃШчвдЯѕЫсЯЫЮЌЫиБЁФЄЮЊжЇГжЮяЃЌЕчгОНсЙћЪЧЃКІСЃжЌЕААзгОЖЏзюПьЃЌЯрЕБгкІС1ЃЧђЕААзЕФЮЛжУЃЛЧАІТжЌЕААзДЮжЎЃЌЯрЕБгкІС2ЃЧђЕААзЮЛжУЃЛІТЃжЌЕААзгОЖЏдкЧАЃІТжЎКѓЃЌЯрЕБгкІТЃЧђЕААзЕФЮЛжУЃЛШщУгЮЂСЃЭЃСєдкЕубљЕФЮЛжУЩЯ(МћЭМ5-3)ЁЃ
ЁЁЁЁ(Жў)ГЌЫйРыаФЗЈ
ЁЁЁЁБОЗЈвРОнВЛЭЌжЌЕААзжаЕААзжЪжЌРрГЩЗжЫљеМБШР§ВЛЭЌЃЌвђЖјЗжзгУмЖШВЛЭЌ(ИЪгЭШ§ѕЅКЌСПЖрепУмЖШЕЭЃЌЕААзжЪКЌСПЖрЕФЗжзгУмЖШИп)ЃЌдквЛЖЈРыаФСІзїгУЯТЃЌЗжзгГСНЕЫйЖШЛђЦЏИЁТЪВЛЭЌЃЌНЋжЌЕААзЗжЮЊЫФРрЃЌМДШщУгЮЂСЃ(chylomicrons)ЁЂМЋЕЭУмЖШжЌЕААз(very low density lipoproteinЃЌVLDL)ЁЂЕЭУмЖШжЌЕААз(low density lipoproteinЃЌLDL)КЭИпУмЖШжЌЕААз(highdensity lipoproteinЃЌHDL)ЃЛЗжБ№ЯрЕБгкЕчгОЗжРыжаЕФШщУгЮЂСЃЁЂЧАІТжЌЕААзЁЂІТжЌЕААзКЭІСжЌЕААзЁЃГ§ЩЯЪіМИРржЌЕААзвдЭтЃЌЛЙгавЛжжжаМфУмЖШжЌЕААз(intermediate density lipoproteinЃЌIDL)ЦфУмЖШЮЛгкVLDLгыLDLжЎМфЃЌетЪЧVLDLДњаЛЕФжаМфВњЮяЁЃHDLдкДњаЛЙ§ГЬжаЗжзгжаЕААзгыжЌРрГЩЗжгаБфЛЏЃЌПЩНЋHDLдйЗжЮЊHDL1ЁЂHDL2гыHDL3ЁЃHDL1ЪЧдкИпЕЈЙЬДМЩХЪГЪБВХГіЯжЃЌHDL2ЮЊГЩЪьЕФHDLЃЌHDL3ЮЊаТЩњЕФHDLЃЌЦфЗжзгжаЕААзГЩЗжЖрЁЃ
ЁЁЁЁбЊНЌжаЕФгЮРыжаЖЬСДжЌЗОЫсПЩгыбЊНЌАзЕААзНсКЯЖјБЛдЫЪфЃЌГЦжЎЮЊжЌЫсАзЕААзЁЃгЩгкжЌРрШОЩЋЪБжЌЗОЫсВЛзХЩЋЃЌЫљвдВЛвзЙлВьЃЌЪЕМЪЩЯЫќЕФЮЛжУгыАзЕААзЯрЕБЁЃ
ЁЁЁЁЖўЁЂбЊНЌжЌЕААзЕФзщГЩ
ЁЁЁЁ(вЛ)жЌЕААзжажЌРрЕФзщГЩЬиЕуЃК
ЁЁЁЁГ§жЌЫсАзЕААзЭтЃЌИїРржЌЕААзОљКЌгаИЪгЭШ§ѕЅЁЂСзжЌЁЂЕЈЙЬДММАЦфѕЅЁЃЕЋзщГЩБШР§гаКмДѓВювьЃЌЦфжаИЪгЭШ§ѕЅдкШщУгЮЂСЃжаКЌСПЮЊзюИпЃЌДяЦфЛЏбЇзщГЩЕФ90%зѓгвЁЃСзжЌКЌСПвдHDLЮЊзюИпЃЌДя40%вдЩЯЁЃЕЈЙЬДММАЦфѕЅвдLDLжазюЖрЃЌМИКѕеМЦфКЌСП50%ЁЃVLDLжавдИЪгЭШ§ѕЅКЌСПЮЊзюЖрЃЌДя60%(МћБэ5-2)ЁЃЊЄ
Бэ5-2ЁЁбЊНЌжЌЕААзЕФзщГЩ
зщГЩ |
| ЗжРр |
УмЖШ |
ПХСЃжБОЖ |
S1 |
ЕчгО |
ЕААзжЪ |
змжЌ |
еМзмжЌЕФ% |
КЯГЩ |
ЙІФм |
| ЁЁ |
ЁЁ |
(mm) |
ЁЁ |
ЮЛжУ |
ЁЁ |
ЁЁ |
ИЪгЭ |
|
ЕЈЙЬ |
|
гЮРы |
ВПЮЛ |
|
| ЁЁ |
|
|
ЁЁ |
ЁЁ |
% |
% |
|
СзжЌ |
|
ЕЈЙЬДМ |
|
|
|
| |
|
|
|
|
ЁЁ |
|
Ш§ѕЅ |
ЁЁ |
ДМ ѕЅ |
|
жЌЗОЫс |
|
|
| CM |
ЃМ0.95 |
90-
|
ЃО400 |
дЕу |
1-2 |
98-99 |
88 |
8 |
3 |
1 |
|
аЁГІеГ |
зЊдЫЭтдДадЧщ |
| |
|
1000 |
|
|
ЁЁ |
ЁЁ |
|
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ФЄЯИАћ |
ИЪгЭШ§ѕЅ |
| VLDL |
0.95- |
30- |
20- |
ІС2Чђ |
7-10 |
90-93 |
56 |
20 |
15 |
8 |
1 |
ИЮЯИАћ |
зЊдЫФкдДад |
| ЧАІТжЌЕААз |
1.006 |
90 |
400 |
ЕААз |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ИЪгЭШ§ѕЅ |
| LDL |
1.006- |
20-25 |
2-20 |
ІТЧђ |
21 |
79 |
13 |
28 |
48 |
10 |
1 |
бЊНЌ |
зЊдЫФкдДад |
| ІТжЌЕААз |
1.063 |
ЁЁ |
ЁЁ |
ЕААз |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЕЈЙЬДМ |
| HDL |
1.063- |
12-20 |
ГСНЕ |
ІС1Чђ |
33 |
67 |
16 |
43 |
61 |
10 |
4 |
ИЮГІ |
зЊдЫЭтдД |
| ІСжЌЕААз |
1.125 |
ЁЁ |
ЁЁ |
ЕААз |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
бЊНЌ |
адЕЈЙЬДМ |
| HDL2 |
1.125 |
7.5-10 |
ГСНЕ |
ІС1Чђ |
57 |
43 |
13 |
46 |
29 |
6 |
6 |
ИЮГІ |
зЊдЫФкдДад |
| HDL3 |
1.120 |
ЁЁ |
ЁЁ |
ЕААз |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
ЁЁ |
бЊНЌ |
ЕЈЙЬДМ |
ЁЁЁЁ(Жў)дижЌЕААз(apoprotein,apo)
ЁЁЁЁжЌЕААзжагыжЌРрНсКЯЕФЕААзжЪГЦЮЊдижЌЕААзЃЌдижЌЕААздкИЮдрКЭаЁГІеГФЄЯИАћжаКЯГЩЁЃФПЧАвбЗЂЯжСЫЪЎМИжждижЌЕААзЃЌНсЙЙгыЙІФмбаОПБШНЯЧхГўЕФгаapoAЁЂapoBЁЂapoCЁЂapoDгыapoEЮхРрЁЃУПвЛРржЌЕААзгжПЩЗжЮЊВЛЭЌЕФбЧРрЃЌШчapoBЗжЮЊB100КЭB48ЃЛapoCЗжЮЊCЂёЁЂCЂђЁЂCЂѓЕШЁЃдижЌЕААздкЗжзгНсЙЙЩЯОпгавЛЖЈЬиЕуЃЌЭљЭљКЌгаНЯЖрЕФЫЋадІСЃТна§НсЙЙЃЌБэЯжГіСНУцадЃЌЗжзгЕФвЛВрМЋадНЯИпПЩгыЫЎШмМСМАСзжЌЛђЕЈЙЬДММЋадЧјНсКЯЃЌЙЙГЩжЌЕААзЕФЧзЫЎУцЃЌЗжзгЕФСэвЛВрМЋадНЯЕЭПЩгыЗЧМЋадЕФжЌРрНсКЯЃЌЙЙГЩжЌЕААзЕФЪшЫЎКЫаФЧјЁЃ
ЁЁЁЁдижЌЕААзЕФжївЊЙІФмЪЧЮШЖЈбЊНЌжЌЕААзНсЙЙЃЌзїЮЊжЌРрЕФдЫЪфдиЬхЁЃГ§ДЫвдЭтгааЉжЌЕААзЛЙПЩзїЮЊУИЕФМЄЛюМСЃКШчapoAIМЄЛюТбСзжЌЕЈЙЬДМжЌѕЃзЊвЦУИ(lecithinЊВcholesterol transferase.LCAT),apoCЂђПЩМЄЛюжЌЕААзжЌЗОУИ(lipoproteinlipase,LPL)ЁЃгааЉжЌЕААзвВПЩзїЮЊЯИАћФЄЪмЬхЕФХфЬхЃКШчapo BЃ48ЃЌapoEВЮгыИЮЯИАћЖдCMЕФЪЖБ№ЃЌapoB-100ПЩБЛИїжжзщжЏЯИАћБэУцLDLЪмЬхЫљЪЖБ№ЕШ(МћБэ5Ѓ3)ЁЃЊЄ
Бэ5Ѓ3ЁЁбЊНЌжЌЕААзЕФдижЌЕААз
| дижЌЕААз |
жЌЕАел |
АБЛљЫсЪ§ |
ЗжзгСП |
РДдД |
ЙІФм |
КЌСП |
| AЂё |
HDLЃЌCM |
243 |
28.300 |
ИЮЁЂГІ |
МЄЛюLCATЃЛHDLЪмЬхХфЛљ |
121.8ЁР13.7* |
| AЂђ |
HDLЃЌCM |
77 |
17.500 |
ИЮЁЂГІ |
ЮШЖЈHDLНсЙЙЃПвжжЦLCATЃП |
33.0ЁР5.0* |
| AЂє |
CMЃЌHDL |
371 |
46.000 |
ГІ |
ЙІФмВЛЧхЃЈДйНјCMКЯГЩЃПЃЉ |
17ЁР2Ёї |
| B-100 |
LDLЃЌVLDLЃЌIDL |
4536 |
512,723 |
ИЮ |
LDLЪмЬхХфЛљЃЈЪЖБ№LDLЪмЬхЃЉ |
87.3ЁР143* |
| B-48 |
CMЃЌCMВаСЃ |
ЁЁ |
265 000 |
ГІ |
ДйНјCMКЯГЩ |
5Ёї |
| CЂё |
VLDLЃЌHDLЃЌCM |
57 |
6,500 |
ИЮ |
МЄЛюLCATЃП |
7.8ЁР2.4* |
| CЂђ |
VLDLЃЌHDLЃЌCM |
79 |
8,800 |
ИЮ |
МЄЛюИЮЭтLPL |
5.0ЁР1.8* |
| CЂѓ |
VLDLЃЌHDLЃЌCM |
79 |
8,900 |
ИЮ |
вжжЦLPLЃЌвжжЦИЮapoEЪмЬх |
11.8ЁР3.6* |
| D |
HDL |
ЁЁ |
22,100 |
ИЮЃП |
ЙІФмВЛЧхЃЈзЊДядЫЕЈЙЬДМѕЅЃП |
10ЁР4Ёї |
| ЁЁ |
VLDLЃЌHDLЃЌCM |
ЁЁ |
ЁЁ |
ЁЁ |
LDLЪмЬхХфЛљЃЌИЮCMВаЪмЬхЃЉ |
ЁЁ |
| E |
CMВаСЃ |
299 |
34,000 |
ИЮ |
ЃЈapoEЪмЬхЃЉХфЛљ |
3.5ЁР1.2* |
ЁЁЁЁ*ЛЊЮївНПЦДѓбЇЩњЛЏНЬбаЪвЖд625Р§ГЩЖМЕиЧје§ГЃГЩШЫВтЖЈЕФНсЙћЁЃЁїЙњЭтБЈЕМВЮПМжЕЁЃ
ЁЁЁЁLCATЃКТбСзжЌЕЈЙЬДМжЌѕЃзЊвЦУИЁЁLPLЃКжЌЕААзжЌЗОУИЊЄ
ЁЁЁЁ(Ш§)жЌЕААзЕФДњаЛ
ЁЁЁЁ1.ШщУгЮЂСЃ(CM)
ЁЁЁЁШщУгЮЂСЃЪЧдкаЁГІеГФЄЯИАћжаЩњГЩЕФЃЌЪГЮяжаЕФжЌРрдкЯИАћЛЌУцФкжЪЭјЩЯОдйѕЅЛЏКѓгыДжУцФкжЪЭјЩЯКЯГЩЕФдижЌЕААзЙЙГЩаТЩњЕФ(nascent)ШщУгЮЂСЃ(АќРЈИЪгЭШ§ѕЅЁЂЕЈЙЬДМѕЅКЭСзжЌвдМАpoB48)ЃЌОИпЖћЛљИДКЯЬхЗжУкЕНЯИАћЭтЃЌНјШыСмАЭбЛЗзюжеНјШыбЊвКЁЃ
ЁЁЁЁаТЩњШщУгЮЂСЃШыбЊКѓЃЌНгЪмРДздHDLЕФapoCКЭapoEЃЌЭЌЪБЪЇШЅВПЗжapoAЃЌБЛаоЪЮГЩЮЊГЩЪьЕФШщУгЮЂСЃЁЃГЩЪьЗжзгЩЯЕФapoCЂђПЩМЄЛюжЌЕААзжЌЗОУИ(LPL)ДпЛЏШщУгЮЂСЃжаИЪгЭШ§ѕЅЫЎНтЮЊИЪгЭКЭжЌЗОЁЃДЫУИДцдкгкжЌЗОзщжЏЁЂаФКЭМЁШтзщжЏЕФУЋЯИбЊЙмФкЦЄЯИАћЭтБэУцЩЯЁЃжЌЗОЫсПЩБЛЩЯЪізщжЏЩуШЁЖјРћгУЃЌИЪгЭПЩНјШыИЮдргУгкЬЧвьЩњЁЃЭЈЙ§LPLЕФзїгУЃЌШщУгЮЂСЃжаЕФИЪгЭШ§ѕЅДѓВПЗжБЛЫЎНтРћгУЃЌЭЌЪБapoAЁЂapoCЁЂЕЈЙЬДМКЭСзжЌзЊвЦЕНHDLЩЯЃЌCMж№НЅБфаЁЃЌГЩЮЊвдКЌЕЈЙЬДМѕЅЮЊжїЕФШщУгЮЂСЃВагрПХСЃ(remnant)ЁЃИЮЯИАћФЄЩЯЕФapoEЪмЬхПЩЪЖБ№CMВагрПХСЃЃЌНЋЦфЭЬЪЩШыИЮЯИАћЃЌгыЯИАћШмУИЬхШкКЯЃЌдижЌЕААзБЛЫЎНтЮЊАБЛљЫсЃЌЕЈЙЬДМѕЅЗжНтЮЊЕЈЙЬДМКЭжЌЗОЫсЃЌНјЖјПЩБЛИЮдрРћгУЛђЗжНтЃЌЭъГЩзюжеДњаЛ(ЭМ5-4)ЁЃ
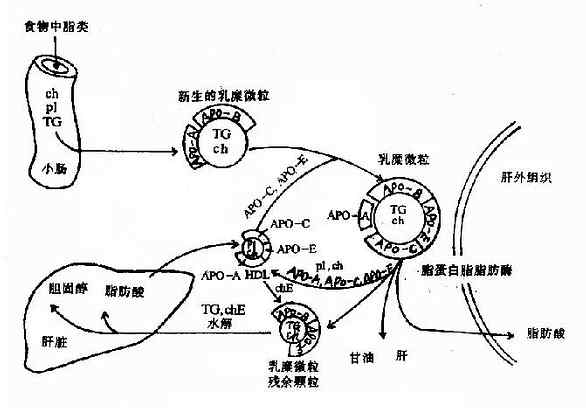
ЭМ5-4ЁЁШщУгЮЂСЃ(CM)ЕФДњаЛЙ§ГЬ
ЁЁЁЁTGЃКИЪгЭШ§ѕЅЃЛpLЃКСзжЌЃЛChЃКЕЈЙЬДМЃЛChEЃКЕЈЙЬДМѕЅЃЛApOЃКдижЌЕААзЃЛHDLЃКИпУмЖШжЌЕААзЁЃ
ЁЁЁЁгЩДЫПЩМћЃЌCMДњаЛЕФжївЊЙІФмОЭЪЧНЋЭтдДадИЪгЭШ§ѕЅзЊдЫжСжЌЗОЁЂаФКЭМЁШтЕШИЮЭтзщжЏЖјРћгУЃЌЭЌЪБНЋЪГЮяжаЭтдДадЕЈЙЬДМзЊдЫжСИЮдрЁЃ
ЁЁЁЁ2.МЋЕЭУмЖШжЌЕААз(VLDL)
ЁЁЁЁVLDLжївЊдкИЮдрФкЩњГЩЃЌVLDLжївЊГЩЗжЪЧИЮЯИАћРћгУЬЧКЭжЌЗОЫс(РДзджЌЖЏдБЛђШщУгЮЂСЃВагрПХСЃ)здЩэКЯГЩЕФИЪгЭШ§ѕЅЃЌгыИЮЯИАћКЯГЩЕФдижЌЕААзapoB100ЁЂapoAIКЭapoEЕШМгЩЯЩйСПСзжЌКЭЕЈЙЬДММАЦфѕЅЁЃаЁГІеГФЄЯИАћвВФмЩњГЩЩйСПVLDLЁЃ
ЁЁЁЁVLDLЗжУкШыбЊКѓЃЌвВНгЪмРДздHDLЕФapoCКЭapoEЃКapoCЂђМЄЛюLPLЃЌДпЛЏИЪгЭШ§ѕЅЫЎНтЃЌВњЮяБЛИЮЭтзщжЏРћгУЁЃЭЌЪБVLDLгыHDLжЎМфНјааЮяжЪНЛЛЛЃЌвЛЗНУцЪЧНЋapoCКЭapoEЕШдкСНепжЎМфзЊвЦЃЌСэвЛЗНУцЪЧдкЕЈЙЬДМѕЅзЊвЦЕААз(cholesteryl ester transfer protein)ажњЯТЃЌНЋVLDLЕФСзжЌЁЂЕЈЙЬДМЕШзЊвЦжСHDLЃЌНЋHDLЕФЕЈЙЬДМѕЅзЊжСVLDLЃЌетбљVLDLзЊБфЮЊжаМфУмЖШжЌЕААз(IDL)ЁЃIDLгаСНЬѕШЅТЗЃКвЛЪЧПЩЭЈЙ§ИЮЯИАћФЄЩЯЕФapoEЪмЬхЖјБЛЭЬЪЩРћгУЃЌСэЭтЛЙПЩНјвЛВНШыБЛЫЎНтЩњГЩLDL(ЭМ5-5)ЁЃ
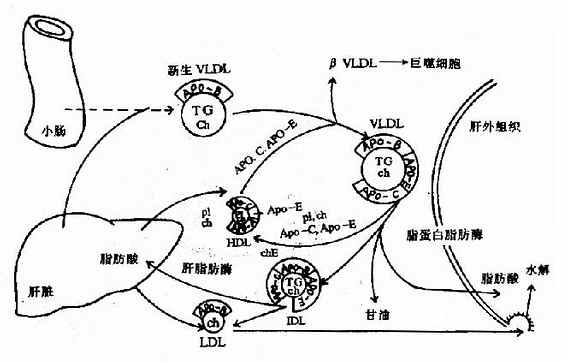
ЭМ5-5ЁЁМЋЕЭУмЖШжЌЕААз(VLDL)ЕФДњаЛЙ§ГЬЊЄ
IDLЃКжаМфУмЖШжЌЕААзЃЛLDLЃКЕЭУмЖШжЌЕААзЃЛ
ЁЁЁЁгЩДЫПЩМћЃЌVLDLЪЧЬхФкзЊдЫФкдДадИЪгЭШ§ѕЅЕФжївЊЗНЪНЁЃ
ЁЁЁЁ3.ЕЭУмЖШжЌЕААз(LDL)
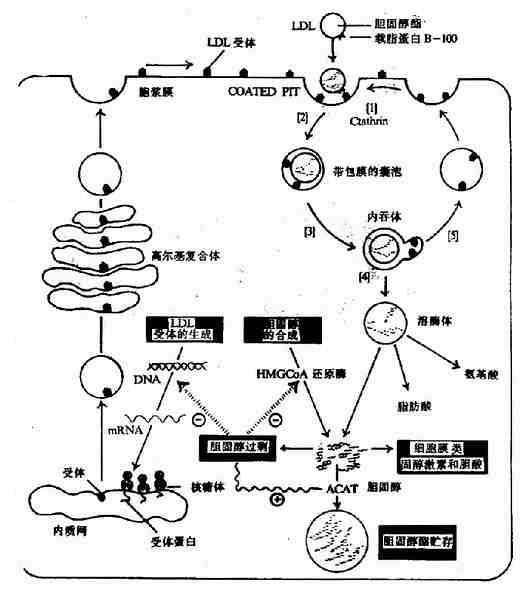
ЭМ5-6ЁЁЯИАћЖдLDLЕФЩуШЁКЭНЕНт
ЁЁЁЁLDLгЩVLDLзЊБфЖјРДЃЌLDLжажївЊжЌРрЪЧЕЈЙЬДММАЦфѕЅЃЌдижЌЕААзЮЊapoB100ЁЃ
ЁЁЁЁLDLдкбЊжаПЩБЛИЮМАИЮЭтзщжЏЯИАћБэУцДцдкЕФapoB100ЪмЬхЪЖБ№ЃЌЭЈЙ§ДЫЪмЬхНщЕМЃЌЭЬШыЯИАћФкЃЌгыШмУИЬхШкКЯЃЌЕЈЙЬДМѕЅЫЎНтЮЊЕЈЙЬДММАжЌЗОЫсЁЃетжжЕЈЙЬДМГ§ПЩВЮгыЯИАћЩњЮяФЄЕФЩњГЩжЎЭтЃЌЛЙЖдЯИАћФкЕЈЙЬДМЕФДњаЛОпгаживЊЕФЕїНкзїгУЃКЂйЭЈЙ§вжжЦHMGЊВCoAЛЙдУИ(HMGЊВCoa reductase)ЛюадЃЌМѕЩйЯИАћФкЕЈЙЬДМЕФКЯГЩЃЛЂкМЄЛюжЌѕЃCoAЕЈЙЬДМѕЅѕЃзЊвЦУИ(acyl CoA:cholesterol acyltransferaseЃЌACAT)ЪЙЕЈЙЬДМЩњГЩЕЈЙЬДМѕЅЖјжќДцЃЛЂлвжжЦLDLЪмЬхЕААзЛљвђЕФзЊТМЃЌМѕЩйLDLЪмЬхЕААзЕФКЯГЩЃЌНЕЕЭЯИАћЖдLDLЕФЩуШЁ(ЭМ5-6)ЁЃ
ЁЁЁЁГ§ЩЯЪігаЪмЬхНщЕМЕФLDLДњаЛЭООЖЭтЃЌЬхФкФкЦЄЭјзДЯЕЭГЕФЭЬЪЩЯИАћвВПЩЩуШЁLDL(ЖрЮЊОЙ§ЛЏбЇаоЪЮЕФLDL)ЃЌДЫЭООЖЩњГЩЕФЕЈЙЬДМВЛОпгаЩЯЪіЕїНкзїгУЁЃвђДЫЙ§СПЕФЩуШЁLDLПЩЕМжТЭЬЪЩЯИАћПеХнЛЏЁЃ
ЁЁЁЁДгвдЩЯПЩвдПДГіЃЌLDLДњаЛЕФЙІФмЪЧНЋИЮдрКЯГЩЕФФкдДадЕЈЙЬДМдЫЕНИЮЭтзщжЏЃЌБЃжЄзщжЏЯИАћЖдЕЈЙЬДМЕФашЧѓЁЃ
ЁЁЁЁ4.ИпУмЖШжЌЕААз(HDL)
ЁЁЁЁHDLдкИЮдрКЭаЁГІжаЩњГЩЁЃHDLжаЕФдижЌЕААзКЌСПКмЖрЃЌАќРЈapoAЁЂapoCЁЂapoDКЭapoEЕШЃЌжЌРрвдСзжЌЮЊжїЁЃ
ЁЁЁЁHDLЗжУкШыбЊКѓЃЌаТЩњЕФHDLЮЊHDL3ЃЌвЛЗНУцПЩзїЮЊдижЌЕААзЙЉЬхНЋapoCКЭapoEЕШзЊвЦЕНаТЩњЕФCMКЭVLDLЩЯЃЌЭЌЪБдкCMКЭVLDLДњаЛЙ§ГЬжадйНЋдижЌЕААздЫЛиЕНHDLЩЯЃЌВЛЖЯгыCMКЭVLDLНјаадижЌЕААзЕФБфЛЛЁЃСэвЛЗНУцHDLПЩЩуШЁбЊжаИЮЭтЯИАћЪЭЗХЕФгЮРыЕЈЙЬДМЃЌОТбСзжЌЕЈЙЬДМѕЅѕЃзЊвЦУИ(LCAT)ДпЛЏЃЌЩњГЩЕЈЙЬДМѕЅЁЃДЫУИдкИЮдржаКЯГЩЃЌЗжУкШыбЊКѓЗЂЛгЛюадЃЌПЩБЛHDLжаapoAIМЄЛюЃЌЩњГЩЕФЕЈЙЬДМѕЅвЛВПЗжПЩзЊвЦЕНVLDLЁЃЭЈЙ§ЩЯЪіЙ§ГЬЃЌHDLУмЖШНЕЕЭзЊБфЮЊHDL2ЁЃHDL2зюжеБЛИЮдрЩуШЁЖјНЕНт(ЭМ5-7)ЁЃ
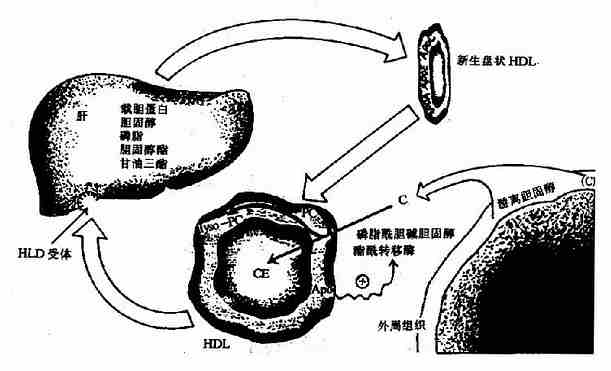
ЭМ5-7ЁЁHDLДњаЛЁЁPCБэЪОСзжЌѕЃЕЈМюЃЌLysoЊВPCБэЪОШмНтадСзжЌѕЃЕЈМю
ЁЁЁЁгЩДЫПЩМћЃЌHDLЕФжївЊЙІФмЪЧНЋИЮЭтЯИАћЪЭЗХЕФЕЈЙЬДМзЊдЫЕНИЮдрЃЌетбљПЩвдЗРжЙЕЈЙЬДМдкбЊжаОлЛ§ЃЌЗРжЙЖЏТіжрбљгВЛЏЃЌбЊжаHDL2ЕФХЈЖШгыЙкзДЖЏТіжрбљгВЛЏГЪИКЯрЙиЁЃ
ЁЁЁЁ(ЫФ)ЁЂИпжЌЕААзбЊжЂ
ЁЁЁЁбЊНЌжЌЕААзДњаЛЮЩТвПЩвдБэЯжЮЊИпжЌЕААзбЊжЂКЭЕЭжЌЕААзбЊжЂЃЌКѓепНЯЮЊЩйМћЃЌЯжжЛНщЩмИпжЌЕААзбЊжЂЁЃ
ЁЁЁЁИпжЌЕААзбЊжЂ(hyperlipoproteinemia)врГЦИпжЌбЊжЂ(hyperlipidemia)ЃЌвђЪЕМЪЩЯСНепОљЯЕбЊжажЌЕААзКЯГЩгыЧхГ§ЮЩТвЫљжТЁЃетРрВЁжЂПЩвдЪЧвХДЋадЕФЃЌвВПЩФмЪЧЦфЫћдвђв§Ц№ЕФЃЌБэЯжЮЊбЊНЌжЌЕААзвьГЃЁЂбЊжЌдіИпЕШЃЌЯжНЋЦфСљжжжївЊРраЭСагкБэ5-4ЁЃЊЄ
Бэ5-4ЁЁИпжЌЕААзбЊжЂЕФРраЭ
| РраЭ |
жЌЕААзБфЛЏ |
бЊжЌЕФБфЛЏ |
ЁЁ |
| ЁЁ |
ЁЁ |
жївЊЩ§ИпЕФжЌРр |
ДЮвЊЩ§ИпЕФжЌРр |
ВЁвђ |
| Ђё |
CMдіИп |
ИЪгЭШ§ѕЅ |
ЕЈЙЬѕЅ |
LPLЛђapoCЂђвХДЋШБЯн |
| Ђђa |
LDLдіИп |
ЕЈЙЬДМ |
ЁЁ |
LDLЪмЬхЕФКЯГЩЛђЙІФмЕФвХДЋШБЯн |
| Ђђb |
LDL VLDLдіИп |
ИЪгЭШ§ѕЅ |
ЕЈЙЬДМ |
вХДЋвђЫигАВЛДѓЃЌжївЊЪмЩХЪГгАЯь |
| Ђѓ |
LDLдіИп |
ИЪгЭШ§ѕЅЕЈЙЬДМ |
ЁЁ |
apoEвьГЃгкШХСЫCMМАVLDLВаСЃЕФЮќЪе |
| Ђє |
VLDLдіИп |
ИЪгЭШ§ѕЅ |
ЕЈЙЬДМ |
ЗжзгШБЯнВЛЧхЃЌЖргЩгкЗЪХжЃЌвћОЦЙ§СПЛђЬЧФђВЁЫљжТ |
| Ђѕ |
CM VLDLдіИп |
ИЪгЭШ§ѕЅ |
ЕЈЙЬДМ |
ЪЕМЪЮЊаЭЂёКЭЂєаЭЕФЛьКЯжЂ |