第四节 糖原的合成与分解
糖原是由多个葡萄糖组成的带分枝的大分子多糖(图4-14),分子量一般在106-107道尔顿,可高达108道尔顿,是体内糖的贮存形式,分子中葡萄糖主要以α-1,4-糖苷键相连形成直链,其中部分以α-1,6-糖苷键相连构成枝链,糖原主要贮存在肌肉和肝脏中,肌肉中糖原约占肌肉总重量的1-2%约为400克,肝脏中糖原占总量6-8%约为100克。肌糖原分解为肌肉自身收缩供给能量,肝糖原分解主要维持血糖浓度(图4-15)。
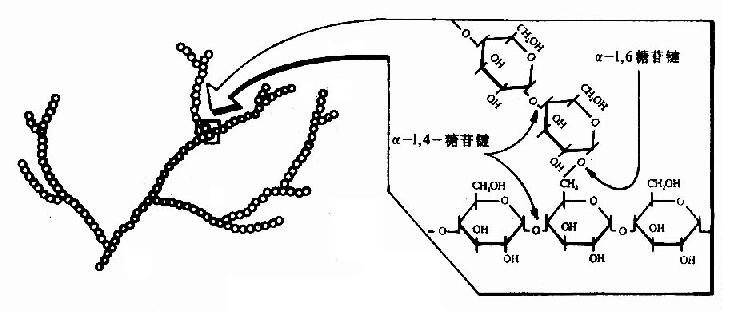
图4-14 糖原的结构
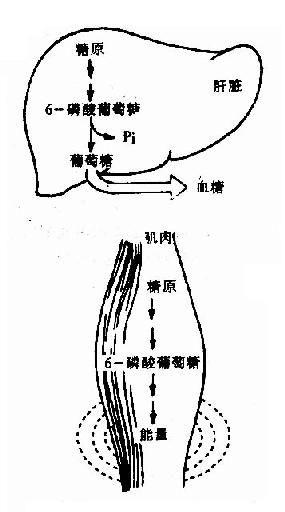
图4-15 肌糖原和肝糖原的功能
一、糖原的合成
由葡萄糖(包括少量果糖和半乳糖)合成糖原的过程称为糖原合成,反应在细胞质中进行,需要消耗ATP和UTP,合成反应包括以下几个步骤:
(1)
(2)
(3)1-磷酸葡萄糖+UTP UDPG+PPi(焦磷酸)
UDPG+PPi(焦磷酸)
(4)UDPG+糖原(Gn) UDP+糖原(Gn+1)
UDP+糖原(Gn+1)
糖原合成酶催化的糖原合成反应不能从头开始合成第一个糖分子,需要至少含4个葡萄糖残基的α-1,4-多聚葡萄糖作为引物(primer),在其非还原性末端与UDPG反应,UDPG上的葡萄糖基C1与糖原分子非还原末端C4形成α-1,4-糖苷链,使糖原增加一个葡萄糖单位,UDPG是活泼葡萄糖基的供体,其生成过程中消耗UTP,故糖原合成是耗能过程,糖原合成酶只能促成α-1,4-糖苷键,因此该酶催化反应生成为α-1,4-糖苷键相连构成的直链多糖分子如淀粉。
机体内存在一种特殊蛋白质称为glycogenin,可做为葡萄糖基的受体,从头开始如合成第一个糖原分子的葡萄糖,催化此反应的酶是糖原起始合成酶(glycogen initiaor synthase),进而合成一寡糖链作为引物,再继续由糖原合成酶催化合成糖。同时糖原分枝链的生成需分枝酶(branching enzyme)催化,将5-8个葡萄糖残基寡糖直链转到另一糖原子上以α-1.6-糖苷键相连,生成分枝糖链,在其非还原性末端可继续由糖原合成酶催化进行糖链的延长。多分枝增加糖原水溶性有利于其贮存,同时在糖原分解时可从多个非还原性末端同时开始,提高分解速度(图4-16)。
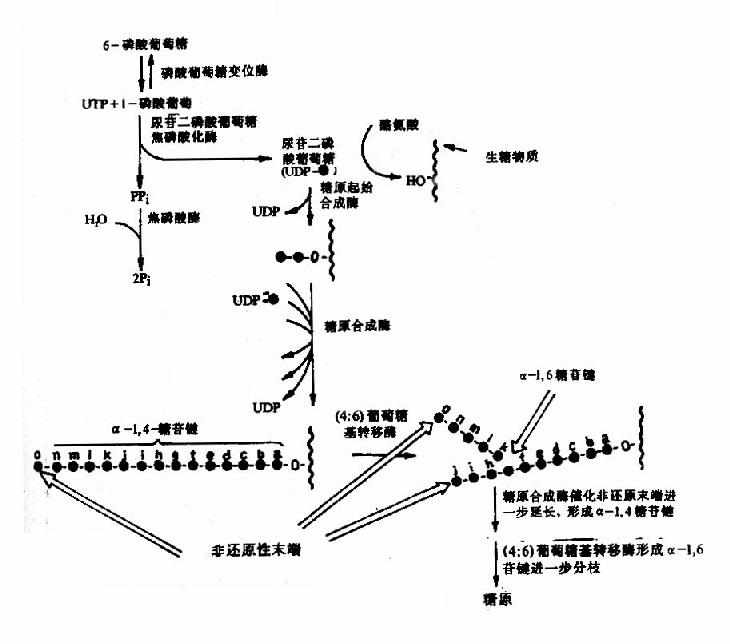
图4-16 糖原合成
二、糖原的分解
糖原分解不是糖原合成的逆反应,除磷酸葡萄糖变位酶外,其它酶均不一样,反应包括:
(1)Gn糖原+Pi G-1-P+g (n-1)
G-1-P+g (n-1)
(2)G-1-P G-6-P
G-6-P
(3)G-6-P+H2O G+PI
G+PI
这样将糖原中1个糖基转变为1分子葡萄糖,但是磷酸化酶只作用于糖原上的α(1→4)糖苷键,并且催化至距α(1→6)糖苷键4个葡萄糖残基时就不再起作用,这时就要有脱枝酶(debranching enzyme)的参与才可将糖原完全分解。脱枝酶是一种双功能酶,它催化糖原脱枝的两个反应,第一种功能是4-α-葡聚糖基转移酶(4-α-D-glucanotrnsferase)活性,即将糖原上四葡聚糖分枝链上的三葡聚糖基转移到酶蛋白上,然后再交给同一糖原分子或相邻糖原分子末端具自由4羟基的葡萄糖残基上,生成α(1→4)糖苷键,结果直链延长3个葡萄糖(图5-6),而α(1→6)分枝处只留下1个葡萄糖残基,在脱枝酶的另一功能,即1,6-葡萄糖苷酶活性催化下,这个葡萄糖基被水解脱下,为游离的葡萄糖,在磷酸化酶与脱枝酶的协同和反复的作用下,糖原可以完全磷酸化和水解(图4-17)。
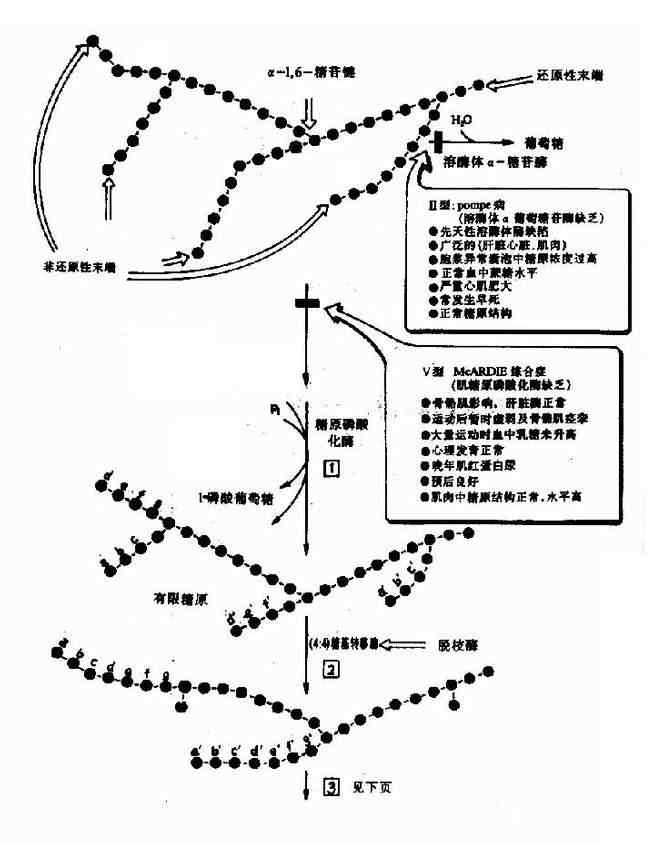
图4-17 糖原的分解

三、糖原代谢的调节
糖原合成酶和磷酸化酶分别是糖原合成与分解代谢中的限速酶,它们均受到变构与共价修饰两重调节。
(一)糖原代谢的别构调节

图4-18 糖原合成和分解的调节
6-磷酸葡萄糖可激活糖原合成酶,刺激糖原合成,同时,抑制糖原磷酸化酶阻止糖原分解,ATP和葡萄糖也是糖原磷酸化酶抑制剂,高浓度AMP可激活无活性的糖原磷酸化酶b使之产生活性,加速糖原分解。Ca2+可激活磷酸化酶激酶进而激活磷酸化酶,促进糖原分解(图4-18)。
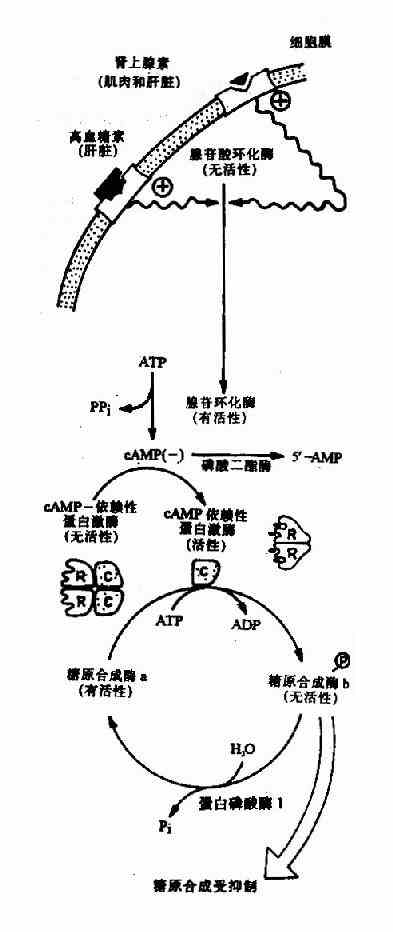
图4-19 糖原合成的抑制
(二)激素的调节
体内肾上腺素和胰高血糖素可通过cAMP连锁酶促反应逐级放大,构成一个调节糖原合成与分解的控制系统。
当机体受到某些因素影响,如血糖浓度下降和剧烈活动时,促进肾上腺素和胰高血糖素分泌增加,这两种激素与肝或肌肉等组织细胞膜受体结合,由G蛋白介导活化腺苷酸环化酶,使cAMP生成增加,cAMP又使cAMP依赖蛋白激酶(cAMp dependent protein kinase)活化,活化的蛋白激酶一方面使有活性的糖原合成酶a磷酸化为无活性的糖原合成酶b(图4?9);另一面使无活性的磷酸化酶激酶磷酸化为有活性的磷酸化酶激酶,活化的磷酸化酶激酶进一步使无活性的糖原磷酸化酶b磷酸化转变为有活性的糖原磷酸化酶a(图4?0),最终结果是抑制糖原生成,促进糖原分解,使肝糖原分解为葡萄糖释放入血,使血糖浓度升高,肌糖原分解用于肌肉收缩。
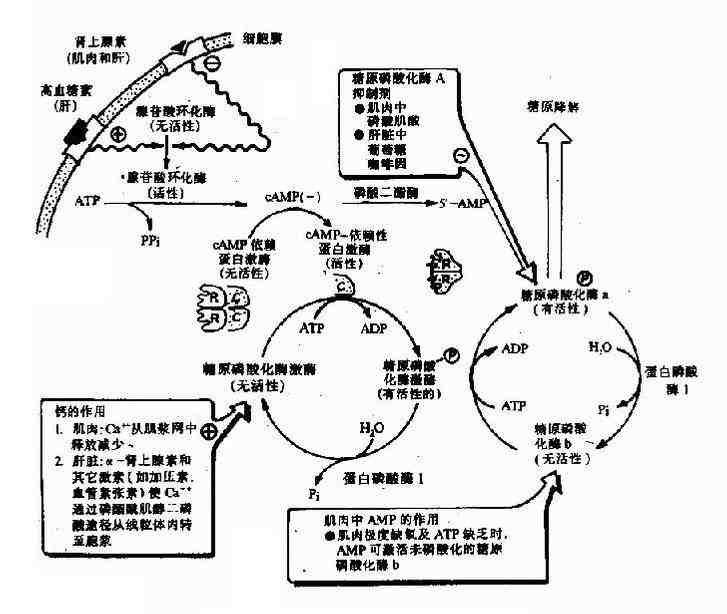
图4-20 糖原合成的抑制