�ڶ��ڡ��ܰ�ϸ��
��������Ӧ���ϸ��������ϸ�������Է�Ϊ�����ࣺ��һ����ָ������Ӧ���������������õ�������ϸ�������ܰ�ϸ�����ڶ�����ָ������Ӧ��������������õĵ��ˣ�����ϸ������������ָ������������ЧӦ����������ϸ������������ϸ��������϶ࡣΪ���ڳ�ѧ�����գ������õ�ϸ����ƺ��������ڱ�4-3��
�����ܰ�ϸ����lymphocyte����������ϵͳ����Ҫϸ���������γɴ�С�ɷ�Ϊ��11��18��m�����У�7��11��m����С��4��7��m�����ࣻ�������ʺ��ܿɷ�ΪTϸ����Bϸ����NKϸ������ͬ���͵��ܰ�ϸ�����Ѵ���̬ѧ�Ϸֱ棬ֻ��ͨ���䲻ͬ�ı����־�Ͳ�ͬ�ķ�Ӧ�Խ������֡�
����һ��Tϸ��
����Tϸ�����������г�����ܰ�ϸ�����ʳ������������ܰ�ϸ����thymus-dependentlymphocyte�������T�ܰ�ϸ����Tϸ������ѪҺ����ѭ���е���Ҫ�ܰ�ϸ����
������һ��Tϸ���ı����־
������Tϸ�������IJ�ͬ�Σ�ϸ������ɱ��ﲻͬ����ķ��ӣ���Щ������ϸ�������йأ�Ҳ����Ϊǩ��Tϸ��˦��״̬�ı����־��
����1��Tϸ�����壨Tcellreceptor��TCR���ֳ�Tϸ����ԭ���壬�ɱ��������г���Tϸ�����棬��Tϸ��ʶ��������ԭ����֮��ϵ��������塣TCR�ɦ��ͦ�����������ɣ�������֮���ɶ�������ӣ���ṹ���ܾ�����IgG���ӵ�һ��Fab�Ρ�������IgG���������ƣ���V��J��C��3������Ƭ������Ļ�����б��룻����������IgG������V����CH1������V��D��J��C��4������Ƭ������Ļ�����б��롣��Tϸ�����������У�������������Ļ�����ͻ������ţ����TCR���и߶ȵĶ�̬�ԣ�����Ӧǧ���Ŀ�ԭ���ӡ�
������Bϸ�������Ig����һ����TCR�������Ե��뿹ԭ��ϡ���Ig���Ӳ�ͬ���ǣ�TCRһ��ֻ�����MHC�������ӵĿ�ԭ����������TCR�뿹ԭ�Ľ�������������ҳ���ֻ�Ц�����������������Ե�ʡ�TCRʶ��ԭ�������ص㹹����MHC�����ԵĻ�����TCR�뿹ԭ��Ϻ���ֱ�ӻTϸ�����������������ڽӵ�CD3������ϸ���ڲ����ݻ��Ϣ��CD4��CD8�����ܹ�Эͬ�ͼ�ǿ�������á�
����һС���ֳ���Tϸ���ı���TCR�����ɦ�/������ɣ������ɦ�/������ɣ���/�������/�����и߶�ͬԴ�ԣ����ҦĻ�������λ�ڦ������λ���ϣ���������Ȼ������ȷ��
����2���طֻ���ԭ��clusterofdifferentiation��CD��Tϸ���ڷֻ���������У���ͬ�ķ����κͲ�ͬ������ܰ�ϸ���ɱ��ﲻͬ�ķֻ���ԭ�����������ܰ�ϸ������Ҫ��־������1986������������֯����ίԱ�Ὠ��Ӧ��CDϵ����ͳһ������ϸ���ֻ���ԭ�������ܰ�ϸ����������ϸ����Ŀǰ�Ѿ�������CD��ԭ70���֣�������ҪCD��ԭ���ڱ�4-1��
��4-1����CD��ԭ����ֲ�������
| ��ԭ |
���� |
�ֲ�ϸ�� |
��Ҫ���ʺ��� |
| CD2 |
T1,E���� |
T,NK |
���CD58,ת���ź� |
| CD3 |
T3,Leu4 |
T |
Tϵ��־,ת���ź� |
| CD4 |
T4,Leu3a |
TH,M�յ�EBVת����B |
MHC�����������,HIV���崫���ź�,TH��־ |
| CD5 |
T1 |
T,����B |
���CD72,����B��־ |
| CD7 |
Leu9 |
T,NK��ǰ�� |
�����ź� |
| CD8 |
T8,Leu2a |
Tc |
I���������,Tc��־ |
| CD10 |
CALLA |
Bǰ�� |
����ø,�����ܰ�ϸ����Ѫ���ı�־ |
| CD11a |
LFA-1�� |
T,B,NK,M |
��ȫ��,�鵼ϸ��ճ�� |
| CD16 |
Fc��R�� |
NK,M��,N |
����IgGFc����,NK��־ |
| CD19 |
B4 |
B |
�����ź� |
| CD21 |
CR2,B2 |
B |
C3d��CD23��EBV���� |
| CD22 |
|
B |
���CD45RO,�����ź� |
| CD23 |
Fc��R�� |
�B,M�յ� |
����IgEFc����,CD21���� |
| CD25 |
IL-2R�� |
�T,B��M |
����IL-2R,L-C���־ |
| CD28 |
|
�TH |
���B7,�鵼Эͬ�̼��ź� |
| CD32 |
Fc��R�� |
B,M��,N,Eo |
����IgGFc����,�����ź� |
| CD35 |
CR1 |
B,M,N,NK |
C3b���� |
| CD40 |
|
B |
���CD40,����TH�����ź� |
| CD45 |
WBC-Ag |
ȫ��WBC |
PTP,����TCR�ź� |
| CD54 |
ICAM-1 |
�L-c |
���LFA-1��CD23 |
| CD56 |
N-CAM |
NK |
NK��־,NKճ������ |
| CD64 |
Fc��R�� |
M,M�� |
������IgGFc���� |
����Tϸ����Ҫ��CD��ԭ�����¼��ࡣ
������1��CD2��������Լ49kD���ǵ��ף�������ȫ����Tϸ����NKϸ�����棻��3�ֿ�ԭ�Բ�ͬ�ķ��ӣ�CD2-1��CD2-2��CD2-3����ɡ�CD2-1��CD2-2�����ھ�ֹϸ�����棬CD2-3�����ڻ��Tϸ�����档Ӧ�ÿ�CD2-2����CD2-3��ֱ�ӻ��ֹ��Tϸ�������dz���Tϸ�������·;����CD2��ճ������֮һ,˳��ԭ�ݳʹ������������á�CD2���ӻ����������ϸ����SRBC����ϣ��ֳ������ϸ�����塣��һ�������£�������Ѫ�ܰ�ϸ����SRBC��ϣ���ѪTϸ���ܽ������SRBC��Ⱦɫ���������¹۲��õ�廨��״���ʳ�E�����γ����飻�ٴ��Ͽ����ڲⶨ����ѪTϸ��������
������2��CD3��Ϊ6�ĸ��Ϸ��ӣ�������ȫ��Tϸ�����棬��Tϸ����ͬ�ı����־��CD3������TCR���ӽ������ӣ���CD3���ӵ�������������IJ��ֱ�TCR���ö࣬����CD3�ɽ�TCR�뿹ԭ����������Ļ�źŴ��ݵ�ϸ���ڲ�������ϸ�������Ӧ�ÿ�CD3����¡����Ҳ��ֱ�ӻTϸ����CD3��TCR�Ĺ�ϵ��ͼ4-5��
������3��CD4/CD8����������������岻ͬ��2�����ӣ���Tϸ����Ⱥ�ı����־������CD4����Ҫ�Ǹ�����Tϸ��������CD8����Ҫ��ϸ������Tϸ����CD4��CD8���ӿ���ǿCD3-TCR��MHC��ԭ��������CD4������ǿ��MHC���ԭ�Ľ�ϣ�CD8��������ǿ��MHC���ԭ�Ľ�ϡ����ٴ�����Ӧ���У�����TCR-CD3��������ԭ-MHC���Ϸ��ӽ�ϵ�������ߣ���ʹϸ�������CD4��CD8���Ӷ�ʧ����ɷ�������Ӧ��
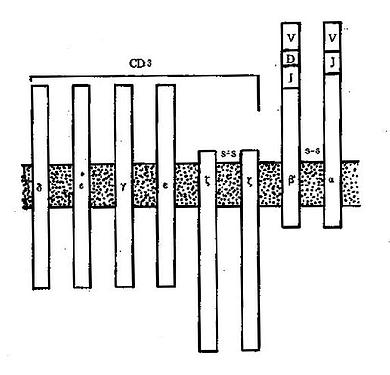
ͼ4-5TCR��CD3�ṹ�������ʾ��ͼ
������4������CD���ӣ�CD7Ҳ��Tϸ���Ĺ�ͬ��־�����ҽ���س�����ϸ�����棬���ɷ�����NKϸ�������������ܰ���ϸ��ǰ���ϡ�ijЩTϸ����Ⱥ��TH������CD28���÷��ӿɴ���ͬ�̼��źţ���ϸ�����ء�
����3�����������־��Ҫ�������¼��ࣺ
������1����֯�����Կ�ԭ��Tϸ����Ҫ����MHC���ԭ��������Tϸ���ɱ���MHC���ԭ��MHC��������������¡�
������2������˿����ԭ���壺����˿����ԭ��mitogen�����˿��ԭ����ͨ����Ӧ����̼���ֹ���ܰ�ϸ��ת��Ϊ�ܰ�ĸϸ����������˿���Ѷ���ֳ��˿��ԭ����ܶ࣬��������ֲ��Ѫ������phytohemagglutinin��PHA����������A��concanavalinA��ConA���ȡ���˿�����PHA��ConA�ȻTϸ����Ҳ�ɽ�˽����ܰ�ϸ��ת���������ж�ϸ�����ߵĹ���״̬��
������3���������壺�ܰ�ϸ�����滹���ڲ������壬�������������������������ȱ�ݲ�����HIV������ȣ�ͨ���������壬��������ѡ���Ե���Ⱦij��Tϸ����Ⱥ������HIV����ͨ��CD4��Ⱦ������Tϸ���������̲���
�������⣬Tϸ���������ж�����ϸ���������壨�������£��������ϸ�����壨CD2���������أ�integrin�����塢ת�����ף�transferrin������ȣ����ж���ճ�����ӣ�����LFA-1��CD2�ȣ���Щ����Tϸ�����һ��������ԡ�
����������Tϸ������Ⱥ���书��
������ȻTϸ���б���TCR��CD2��CD3��CD7�ȹ��ԣ�������һ���Ǿ�һ�Եĸ���Ⱥ�壻���Էֻ�Ϊ���ﲻͬCD���ӡ����в�ͬ�����Ե���Ⱥ��subset�������н�Ϊ��ȷ��������Ⱥ�Ǹ�����Tϸ����ϸ������Tϸ����Tϸ�������ַֻ��Dz�����ġ�
����1��������Tϸ����helpTcell��TH�����ܹ�����Bϸ���ֻ��ɿ������ϸ���ͷŴ�ϸ������Ӧ���һ��ϸ��Ⱥ������CD4��������CD8��TH�����ͷ�ϸ�����������Ե���Tϸ����Bϸ�������ˣ�����ϸ������������ϸ���Ļ��ԡ�TH�����CD4������Ҫ����MHC���������أ��������븨��������أ���˲���CD4+ϸ����ϸ�������ԣ���ijЩCD8+ϸ���и������á�
�������ݲ���ϸ�����ӵIJ�ͬ��TH�ɱ��ֳ�3�ͣ�TH1��TH2��TH0��TH1����IL-2��IFN�ã�������ǿϸ������Ӧ�𣬴ٽ�Bϸ���ϳ�IgM��IgG2�������ϸ����TH2����IL-4��IL-5����ǿIgG1��IgE�ĺϳɣ����Ӿֲ���ѭ������������ϸ����������TH0��������4��ϸ�����ӣ����TH1��TH2��������ԡ���ͬ�Ŀ�ԭ�յ�������Ӧ������ɲ�ͬ���͵�TH���ƣ���������Ⱦ������Ӧ����TH2���ƣ�����IgEѪ��ˮƽ���ߺ���������ϸ���������ӣ���ԭ�棨����ʲ��ԭ�棩��Ⱦ������Ӧ������TH1���ƣ�����ϸ������Ӧ������ԭ��ĸ�Ⱦ��
��������Ӧ��Ŀ���ѡ���TH���ͷֻ��Ļ���Ŀǰ�в��������ϸ�����������еľֲ�������Ӱ��ϸ���ķֻ��������Ѿ�֤��IL-4�ɴٽ�TH2Ӧ�𣬶�IFN�ÿ�����TH2��Ӧ��
����2��ϸ������Tϸ����cytotoxicTcell��Tc��CTL�����������ܽ��ϸ����һ��ϸ����Ⱥ������CD8��������CD4��Tc�ܹ�ɱ�����б���MHC������Ӳ�����ݳʿ�ԭ�İ�ϸ��������ɱ���ǿ�ԭ�����Եġ�NHC������������Եĺ��г�Ч�ġ�Tcϸ����ɱ��һ��ϸ������ת����һ����ϸ����������ʹ����ɱ�˹��ܡ����Tcϸ���ڿ�������Ⱦ�����������ߡ���ֲ�ųⷴӦ��ijЩ�������߲�������Ҫ���á�
����3������Tϸ����Ⱥ����������ѧ��������д��������Ҫ��Tϸ����Ⱥ������������ѧ�����Ľ�������Щ��Ⱥ���ѵõ�֤ʵ������֤���DZ����Ⱥ��
������1��������Tϸ����suppressorTcell��Ts����������Ϊר����ʹ�����������õ�Tϸ����Ⱥ����1960���״α���ijЩ�ܰ�ϸ�����������������Ժ�30�������д���������������Ts�Ļ��ԣ�������û���˷����ֻ�������������õ�������Tϸ��Ⱥ������Ŀǰ��Ϊ����ν������Tϸ�������ܾ���TH1����ΪTH1����������ϸ������Ӧ���⣬�����ڵ�IFN�û�������IgG1�IJ�����
������2���ٷ��ͳ�����Ӧ��Tϸ����delayed-typehypersensitivityTcell��TDTH����������Ϊ������ٷ��ͳ�����Ӧ��Tϸ�����й�������Ⱥ���������������Щ��������֤������ר���յ��ٷ��ͳ�����Ӧ��Tϸ��Ⱥ��ʵ���ϣ��ٷ��ͳ�����Ӧ������TH��Tc������ġ�
������3����/��Tϸ��������TCR�ɦúͦ�����ɡ�����ͨ����/������ϸ��Ⱥ����Ⱥϸ���������٣�������Tϸ��������5�������ǿ�����CD4��/CD8����Ҳ������CD4��/CD8+������ϸ�������ԣ�Ҳ���ͷ�ϸ�����ӣ���Ƥ���д��ڽ϶࣬�������д���һ��֤����
����������Tϸ���ķ�����ֲ�
����Tϸ����Դ�ڹ����ϸ�����������ڷ������졣Ǩ�������ٵij��ڣ���Щ����ϸ��ֻ����Tϸ��ϵ����������CD2��CD7��������CD4��CD8����Ϊ˫��������ϸ�����ڴ��ڼ䣬TCR����ʼ���ţ������ŵĽ�������˦��ͦ���ʱ��TCR��CD3�㿪ʼ��ˮƽ��Эͬ������ϸ�����档��ʱCD4��CD8Ҳ��ʼ�������ͬʱ������ϸ��Ĥ�ϣ�������ϸ����Ϊ˫��������ϸ����
������˫��������ϸ���Ѿ�����TCR��CD3�����ܹ�ʶ��������MHC���ӵĿ�ԭ��������ʱ��TCR�̼������յ�ϸ����ֳ�������յ�ϸ���������������£��������ڴ��ڼ��������Ŀ�ԭͨ�������������ʣ�����������ϸ�������϶���������Ӧϸ�������������Ϊ����ѡ��negativeselection������һ���棬��ʶ���������ԭ��˫����ϸ�����Լ�����������TCR��CD3���ӵı�����ǿ�����Ҷ�ʧCD4��CD8�����е�һ�����ֻ�Ϊֻ����CD4��CD8�ĵ���������ϸ�����������Tϸ��������ѡ���Ϊ����ѡ��positiveselection����Tϸ���������ڵ���Ҫ�����ص��ͼ4-6������������ǣ�����ѡ��ʱ��û��������ԭ��TCR��ϣ�������ѡ��ķ�������Ӧ��ϸ��ȴҲ��ʶ��������MHC��
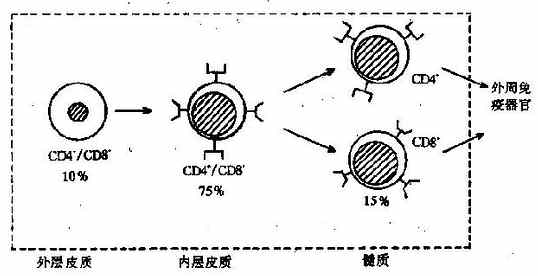
ͼ4-6Tϸ���������ڵķ�������
���������Tϸ���뿪���ٽ���Ѫѭ�����ֲ��������������ٵ������������������ܰͽ�ĸ�Ƥ�����ȣ��ܿ�ԭ�̼����������Ӧ��Tϸ�����ܰ�ϸ����ѭ������Ҫϸ������ѪҺ��Լռ60����70�������ܰͽ�Լռ65����85�������ص�����ռ90�����ϡ������ٷ�����ȫʱ��Tϸ���������裬����Ѫ�ܰ�ϸ���������٣������ܰ����ٵ�����������ή����
��������Bϸ��
����Bϸ���������෨���һ���ͬ�����٣����裩�ڷ��������ϸ������˳�Ϊ�����һ�����������ܰ�ϸ����bursaorbonemarrowdependentlymphocyte�������B�ܰ�ϸ����Bϸ����Bϸ���ܿ�ԭ�̼���ɲ�������Ľ�ϸ����
������һ��Bϸ���ı����־
����1��������������surfaceimmunoglobulin��SLg����Bϸ����������Եı����־������Bϸ��Ĥ�������SIgM��SigD������Bϸ��ֻ����SIgM���ӣ�SIg�������ṹ��Ig��ͬ����SIgM�ǵ�����ӡ�һ��Bϸ��������������SIg���ӣ��������Զ���ϸ�����ڵ�Ig������ͬ������ϸ���ڷֻ������л���ͻ���Ig�������ŵĽ����һ������������������3��106���ɲ�����ͬ������ӵ�ϸ����¡��SIg�Ĺ�����ΪBϸ������Ŀ�ԭ���壬������Ӧ��ԭ�����Խ�ϣ�������ԭ�����㴦������������鵼�Ľ����Bϸ������ԭ����Ҫ��ʽ��
����SIgM��SIgD���ӵ��Ȼ��˲���ϸ��Ĥ����ֻ�м������������ȣ�������TCRһ����SIg�������ܶ�������ϸ���ڴ��ݴ̼��źţ������������������ص��������ֿ�Ĥ�ǵ��ח�Ig-����Ig-�¡������ֵ������������һ�𣬸���һ������Ķ�����ϸ��������˿�����Tϸ�������CD3ЭͬTCRһ������SIg��ϸ���ڴ��ݴ̼��źš�
����2��MHC��CD��ԭ�����Bϸ���������MHC������ӣ����ʹBϸ����Ϊ��ԭ�ݳ�ϸ����������������أ���������£���
����Bϸ���ڷֻ���������пɱ��ﲻͬ��CD���ӣ�����ijЩ����ΪBϸ���ı�־��ijЩ����ϸ��������ء�CD10ֻ������Bǰ��ϸ����CD19��ԭʼ�������Bϸ�������ڣ���CD22ֻ�ڳ���Bϸ��������Ǿ������ı�־��CD21��CD35�Dz������壻CD23��CD32����Ig��Fc���壻CD40����THϸ���ϵ��������ϣ��Ӷ�����TH�ĸ������á�
����3��������־������
������1��Fc���壺Bϸ��������IgG��Fc���壨CD32������Bϸ�������йء�Fc���廹���뿹������ĺ�ϸ�������γ�EACõ�廨�����Ǽ���Bϸ���Ĵ�ͳ����֮һ��
������2���������壺CR�����ڳ���Bϸ���ı��棬CR1����C3b��C4b��ϣ��ٽ�Bϸ��������Ʋ�����CR2���壨CD21������C3d��ϣ�ͬʱҲ��EB���������塣
������3��˿�����壺Bϸ��������˿����ԭ��Ҫ��֬���ǣ�LPS������˿��ԭ���Bϸ��Ҳ���Էֻ���ֳ��
�������⣬Bϸ�����滹�ж���ϸ�����ӣ���IL-1��IL-2��IL-4����IFN�õȣ������壬�벻ͬϸ�����ӵĽ�Ͽ�ʹBϸ��������Ӧ��������ԡ�Bϸ����Tϸ���ıȽϼ���4-2��
��4-2Tϸ����Bϸ����NKϸ������״�Ƚ�
| ��״ |
Tϸ�� |
Bϸ�� |
NKϸ�� |
| �ֻ����첿λ |
���� |
���� |
? |
| �����־ |
|
|
|
| ����ĤIg |
�� |
�� |
�� |
| TCR |
�� |
�� |
�� |
| HLA���ԭ |
������ |
�� |
�� |
| CD2 |
�� |
�� |
�� |
| CD3 |
�� |
�� |
�� |
| CD19��CD20 |
�� |
�� |
�� |
| CD16��CD56 |
�� |
�� |
�� |
| CR |
�� |
�� |
���� |
| FC��R |
�� |
�� |
�� |
| ϸ���ֲ������� |
|
|
|
| ����Ѫ |
60��70 |
20��30 |
10��15 |
| ���� |
��5 |
��95 |
���� |
| �ص��� |
90 |
10 |
�ټ� |
| Ƣ |
30��50 |
50��60 |
�ټ� |
| �ܰͽ� |
65��85 |
15��35 |
�ټ� |
����������Bϸ���ķֻ�����
�����˺Ͳ��鶯��Bϸ���IJ����������ͳ�����ڹ�������ɣ�������̰�����һϵ�еİ��ڻ���ͱ����־�ı仯��ͼ4-7����
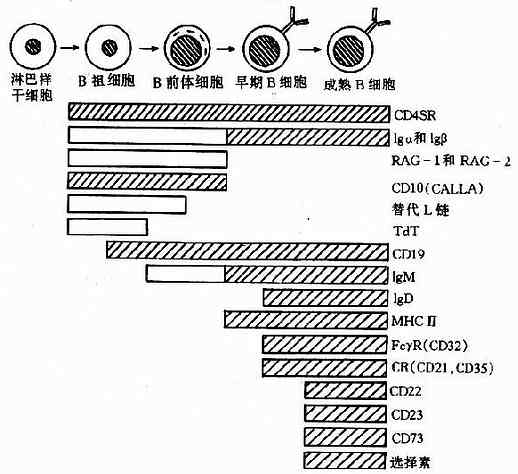
ͼ4-7Bϸ���������̼���������
����B��ϸ����pro-Bcell��ֻ����CD10��CD19��ĩ����������ת��ø��TdT�������ʱ�ڵ���Ҫ�仯�������������ţ����ŵ�ʧ����ռ50�������ɹ������ŵ���ϸ���������ɹ������ſɵ��²���������m������ڴ����������У�������m����δ��������������ﻯ��������surrogatelightchains�����ϣ����γɵ���ʱ��������ת�Ƶ�ϸ��Ĥ�ϣ�����ת�Ʋ������ź�ʹ������������ֹͣ��������ʱ��������ʧ��ϸ������������һ�Ρ�
����Bǰ��ϸ����pre-Bcell���ڽ������ǰ���ٷ�����ֳ����ʱ�ڵ���Ҫ�仯�������������š����ŵĻ�������ʻ�����������е���������γ�4�ĵ�IgM���壬��ת�Ƶ�ϸ��Ĥ�ϡ���һ�ź�ʹϸ����������ڣ������Ե�ʧȥIg�������ŵ�����������ܿ�ԭ�̼�����ֳ�����з��Ӵ�ϸ����������SIgͬ�������ԵĿ�����ӡ�����Bϸ�����������������־������IgD���鳲���塢MHC������ӡ�CD22��CD23��CD40�ȡ�
����������Bϸ������Ⱥ�ͷֲ�
������ͬ��¡�ij���Bϸ�����ﲻͬ�����ԵĿ�ԭ���壬��Ȼ������������֮��IJ�𣬵��Ǻ����ҵ�������������־��������Tϸ����CD4��CD8��������Bϸ���ֳɼ�����Ⱥ������������һС���ֳ���Bϸ������CD5����Щϸ������������Ӧ�������������壩��أ�CD5�������ڼ������������ܰ���Ѫ��ϸ���ϡ�
����δ����Bϸ�����ڹ����ڣ�������������ڹ���2��3�죬�������־������ȫ����뿪�������Ѫѭ�����ֲ��������������ٵķ�������������Bϸ����ѪҺ��Լռ20����30�������ص����в�����10�������ܰͽ���Լռ15����35������Ƣ��������࣬�ɴ�60����Bϸ���ܿ�ԭ�̼�������������������м�����ֳ�ֻ�Ϊ��ϸ�������ڿ��壨��������£���
����������Ȼɱ��ϸ��
������Ȼɱ��ϸ����naturalkiller��NK���ɷ�������ɱ������ϸ���Ͳ�����Ⱦϸ�����ܰ���ϸ����Ŀǰ��NKϸ�����˽��в����������������ѧDZ��ֵ�ý�һ��̽�֡�
������һ��NKϸ����һ������
����NKϸ������ϴ��а�������������̬�϶�����ɫ��NKϸ������û��TCR��CD3��Tϸ����־��Ҳû��SIg��CD40��Bϸ����־�����������Ϊ�ޱ�־ϸ������ϸ��(nullcell)��NKϸ������CD56������Ϊϸ����ϵͳ��־���뵥�ˣ�����ϸ��һ������CD16�ȷ��ӣ�����ɱ�˹�����ء�
����NKϸ��Ҳ��Դ�ڹ���ǰ��ϸ�����䷢���ص�ͷֻ������в���ȷ�����䷢�������ͷֻ���Ȼ��ͬ��Tϸ����Bϸ������ΪT��Bϸ����������ȱ��ʱNKϸ������������T��Bϸ������ʱ����NKϸ��ȱ�ݡ�
����NKϸ�����������٣�������Ѫ��Լռ�ܰ�ϸ��������15������Ƣ��Լ3����4����Ҳ�ɷ����ڷΡ��κͳ�ճĤ���������١��ܰͽ���ص����к�����
����������NKϸ���Ļ��Ժ���
����1����Ȼɱ�˻���NKϸ������Ҫ������ɱ������ϸ���Ͳ�����Ⱦϸ������Tcϸ����ͬ������ɱ�˲���ҪTCRʶ���ϸ���ϵĿ�ԭ��Ҳ����Ҫʶ���ϸ���ϵ�MHC���ӣ���˿����ڰ�ϸ����¶��������ʹɱ�˹��ܣ�����Ҫ���ȵĿ�ԭ���������Գ�Ϊ��Ȼɱ�ˡ�
������Ȼɱ�˵�ʶ������в������һ����Ҫ�Ĺ۵���NKϸ������ɱ����������壺һΪ�̼��ͣ��ɴ����ź�ʹϸ���ͷ�IFN�ú�THF����ϸ�����ӣ��Ӷ�ɱ��ϸ������һΪ�෴���õ������͡���NKϸ��ֻ����̼�������ʱ����ɱ�����á�����֪������������һ�ֳ�ΪLy-49��Ĥ������ӣ�����������MHC������ӣ���ϸ��������������ʢʱ��NKϸ����ʧȥ��Ȼɱ�˵�������
����2��ϸ�����ӻ��ɱ������NKϸ����ɱ�˻��Կ�ͨ��ijЩϸ�����ӣ�����IL-2�����յ���������ǿ��������ϸ����Ϊ�ܰ����ӻ��ɱ��ϸ����lymphokine-activatedkiller��LAK��������Ѫ������ϸ���ں�IL-2���������з�����ɵõ�LAKϸ����NKϸ��������Ҫ�ɷ֡���LAKϸ����������������DZ����һ�������Ʒ���
����3������������ɱ������NKϸ��������IgG��Fc���壨CD16����������ͨ�������ý��NKϸ����ɱ�˿�������İ�ϸ������������Ļ��Գ�Ϊ����������ϸ���鵼��ϸ�������ã�antibody-dependentcell-mediatedcytotoxicity��ADCC��������������£���ϸ����ʶ��ѡ��ȡ���������Կ��塣